
; Forêt vierge au soleil couchant; um 1910")
; Forêt vierge au soleil couchant; um 1910")
Wat is ecologie? (serie 1)

.
Sommige mensen zijn dol op een gladgeschoren gazon. Voor hen geldt: een dicht grasveld is goed, onkruid ertussen is fout. Alles wat geen gras is, bijvoorbeeld weegbree en paardenbloem, geldt als onkruid. (En een krop sla midden op een gazon, dat hoort al helemaal niet.)
De natuur is niet echt hartelijk voor deze mensen, want wie het ook is die een grasveld aanlegt, hij of zij houdt andere planten bijna niet tegen. Voor ons menselijk voortbestaan is een gazon van geen enkel belang. Het is wel een goed onderwerp om over ecologie te praten.
Grasvelden houd je in stand door het gras stelselmatig te maaien. Sommige onkruiden worden verwijderd door ze uit te graven. Ook kun je bestrijdingsmiddelen tegen het onkruid tussen het gras gebruiken. Kale plekken moet je opnieuw inzaaien of beplanten. Anders is er al gauw geen echt gazon meer.
Maaien voorkomt dat gras gaat bloeien en zich uitzaait. Sommige mensen gebruiken maaimachines die het gras opvangen of harken het bij elkaar. Zo verdwijnt het gemaaide gras vaak naar de composthoop. In ecologische termen betekent dit dat een belangrijk deel van het lévende materiaal dat het grasveld produceert, verwijderd wordt. Er is zo een gestadig verlies aan voedingsstoffen uit het milieu van het grasveld. Het kan op den duur nodig zijn om mest te gebruiken. Anders zal het gras onvoldoende groeien.
Bij het maaien wordt het aanwezige onkruid meegemaaid. Sommige soorten onkruid, zoals paardebloemen, reageren op regelmatig maaien door dicht bij de grond te bloeien. Ze bloeien met een veel kortere stengel. In zekere zin veroorzaakt het maaien een vorm van selectie. Het maaien bevoordeelt individuen en soorten die dicht bij de grond bloemen en zaden kunnen vormen. Hierom zijn madeliefjes zo succesvol in een grasveld.
Als het gemaaide gras blijft liggen, komen er dieren als slakken en pissebedden. De snelheid waarmee het gemaaide gras wordt afgebroken, hangt af van de vochtigheid van het grasveld en de heersende temperatuur. Het sproeien van het grasveld schept een milieu dat gunstig is voor regenwormen. Net als slakken en pissebedden breken regenwormen dood materiaal af. Grasvelden voorzien in het levensonderhoud van grote aantallen regenwormen.
Een gazon is dus een toestand waar enkele soorten overheersen en elke natuurlijke tendens naar meer soorten door de tuinier wordt onderdrukt. Grasvelden blijven onder normale omstandigheden niet vanzelf bestaan. Zonder mensen die ze bijhouden, ontstaat er na verloop van tijd een geheel andere samenstelling van planten.
Gazons tonen ons ecologische principes.

.
Een merel ziet een worm zodra er maar een puntje boven de grond uitsteekt. Hij is er goed in om de hele worm uit zijn gang te trekken zonder hem af te breken. Als je dat zelf probeert met een pincet, zal blijken dat dit niet zo eenvoudig is als het lijkt. Ofwel de worm trekt zich snel terug, òf het puntje breekt af wanneer de worm zich vastzet tegen de wand van zijn gang.
Regenwormen leven niet diep onder de grond. Vooral bij nat weer en ’s nachts komen ze boven. Ze voeden zich met dood en rottend plantenmateriaal. Met hun gespierde keelholte nemen ze grote hoeveelheden grond op die humus bevat. Dit wordt door de darm gevoerd, waarna de uitwerpselen verspreid over het gras achtergelaten worden. Wormen zijn zo talrijk en ze verwerken bij hun voeding zoveel grond dat ze tenslotte alles onder hun uitwerpselen bedekken.
Veel dieren eten wormen. Ze vormen een goede bron van eiwitten. De meest efficiënte wormeneters leven onder de grond. Maar als het gras kort gehouden wordt, zijn wormen ook bereikbaar voor vogelsoorten die hun voedsel zoeken op open plaatsen in de bovenste laag van de grond. De zanglijster en de merel zijn zulke vogels.*
Soms heeft een merel, die in het voorjaar bij droog weer zijn jongen moet voeren, tekort aan voedsel omdat de wormen zich niet laten zien. De vogel moet dan naar ander voedsel uitzien, zoals slakken. Als het nat weer is, zijn er genoeg wormen.
Het aantal wormen schijnt nauwelijks onder de jacht van de merels te lijden. De lege plaats van een verdwenen worm wordt kennelijk al gauw ingenomen door een andere worm. Er is een duidelijke wisselwerking tussen de aantallen wormen en de aantallen merels. Beide hebben een groot vermogen zich te vermenigvuldigen. Dat hun aantallen niet exploderen is zo’n elementair verschijnsel, dat we de betekenis ervan vaak over het hoofd zien.
We kunnen concluderen dat een stuk gras een bepaald aantal wormen kan ‘onderhouden’. Wormen voorzien op hun beurt een zeker aantal wormeneters van voedsel. Stellig wordt er een soort evenwicht gehandhaafd. We gaan uitzoeken hoe dat precies zit.
Een gazon biedt allerlei diersoorten een plek om te leven.
.
Het verschil tussen de soorten vogels in de tuin is gemakkelijk te zien. Het is moeilijker de verschillende soorten grassen of wormen te onderscheiden. Wanneer spreken we van een aparte soort?
Voortplanting zien we alleen tussen twee organismen met nagenoeg dezelfde eigenschappen. Tussen andere blijkt het niet te gaan. Tot één soort rekenen we alle organismen waarvan mannetjes en vrouwtjes onderling tot voortplanting in staat zijn. (Overigens zullen we later zien dat het begrip soort ingewikkelder is.)
Mensen vormen één soort omdat ze zich in principe vrij met elkaar kunnen voortplanten. In de praktijk zijn er echter geografische of sociale barrières die vermenging verhinderen. Barrières voor organismen van eenzelfde soort zijn in de natuur een gegeven. Vinken in het Amsterdamse Bos zijn van dezelfde soort als vinken in het Kralingse Bos. Maar ze zullen zich niet met elkaar voortplanten omdat ze te ver van elkaar leven.
Om binnen één soort toch onderscheid te kunnen maken, spreken we van de vinkenpopulatie in het Amsterdamse Bos en de vinkenpopulatie in het Kralingse Bos. Exemplaren van één populatie ‘doen’ het met elkaar.
Het woord populatie komt van het latijnse woord populus, dat volk betekent. Lange tijd gebruikte men het begrip populatie alleen voor groepen ménsen. Ecologen gebruiken het begrip populatie om de groep organismen aan te geven die samen leeft en zich samen voortplant. Het is dus onjuist om over een populatie te spreken als het om méérdere soorten gaat.
Van een onkruid dat spontaan in onze tuin opkomt, zoals zuring, kunnen we zeggen dat alle exemplaren in de tuin tot één populatie behoren. Ze vormen een populatie omdat ze zich in de praktijk samen voortplanten. Hiertoe behoren niet exemplaren van dezelfde soort, die tien jaar later op een ander stuk land gevonden worden. Deze behoren in elk opzicht tot een andere populatie. Natuurlijk zijn de grenzen niet strak te trekken. Ook gelden niet voor elke soort dezelfde grenzen. Slakken, die in een haag leven en door velden en wegen geïsoleerd zijn van andere slakken van dezelfde soort, vormen een aparte populatie. Voor vogels betekenen dezelfde velden en wegen geen barrière. Omdat ze veel mobieler zijn strekken hun populaties zich over grotere gebieden uit.
Ecologie begint met de begrippen soort en populatie.

.
Wie geld wil sparen en de bank niet vertrouwt, kan elk jaar honderd euro in een oude kous stoppen. Ieder jaar groeit dat bedrag dan met een constante hoeveelheid van honderd euro. We noemen dat lineaire groei. Wie de bank wel vertrouwt en zijn geld daarheen brengt, krijgt er rente over (al is dat op het ogenblik niet veel). Laat je dat geld op de bank staan, dan groeit het spaartegoed elk volgend jaar méér dan het jaar ervoor. Dan spreken we over exponentiële groei.
Een bekend raadseltje is dat van de waterlelie in de vijver: Een populatie waterlelies groeit in een vijver en verdubbelt zich iedere dag. Na dertig dagen is de vijver helemaal volgegroeid. De vraag is: Wanneer was de vijver nog maar half vol? Het antwoord: op de 29e dag, de dag vóór de waterlelies de hele vijveroppervlakte bedekken. Dit raadsel laat zien dat een proces van exponentiële groei eerst onbelangrijk kan lijken, terwijl de gevolgen een ogenblik later al zorgwekkend kunnen worden.
Gist gebruikt voor zijn groei suiker. De minuscule gistcellen groeien in aantal door zich elke paar minuten te delen. In korte tijd kunnen een paar cellen uitgroeien tot een populatie van miljoenen. Als er een onbeperkte hoeveelheid suiker is en de omstandigheden gunstig zijn voor het overleven van de gistcellen, nemen de cellen exponentieel in aantal toe. Een populatie die met één cel begint, zou zich als volgt ontwikkelen: 1, 2, 4, 8, 16, 32, 64, 128, 256, 512, 1024, 2048, 4096, 8192… enz.
Binnen korte tijd zullen er zo ontelbaar veel gistcellen zijn. In de praktijk is de voorraad suiker natuurlijk altijd beperkt. De suiker raakt op en de groei van de populatie stopt dan door gebrek aan voedsel. Ook kan de groei geremd worden door gebrek aan ruimte of doordat de omgeving vergiftigd raakt met de stofwisselingsproducten (de ‘poep’ en de ‘pies’) van die enorme groei aan gistcellen. De groei ervan lijkt nog dramatischer als de aantallen worden uitgezet in een grafiek, zoals in figuur 1.
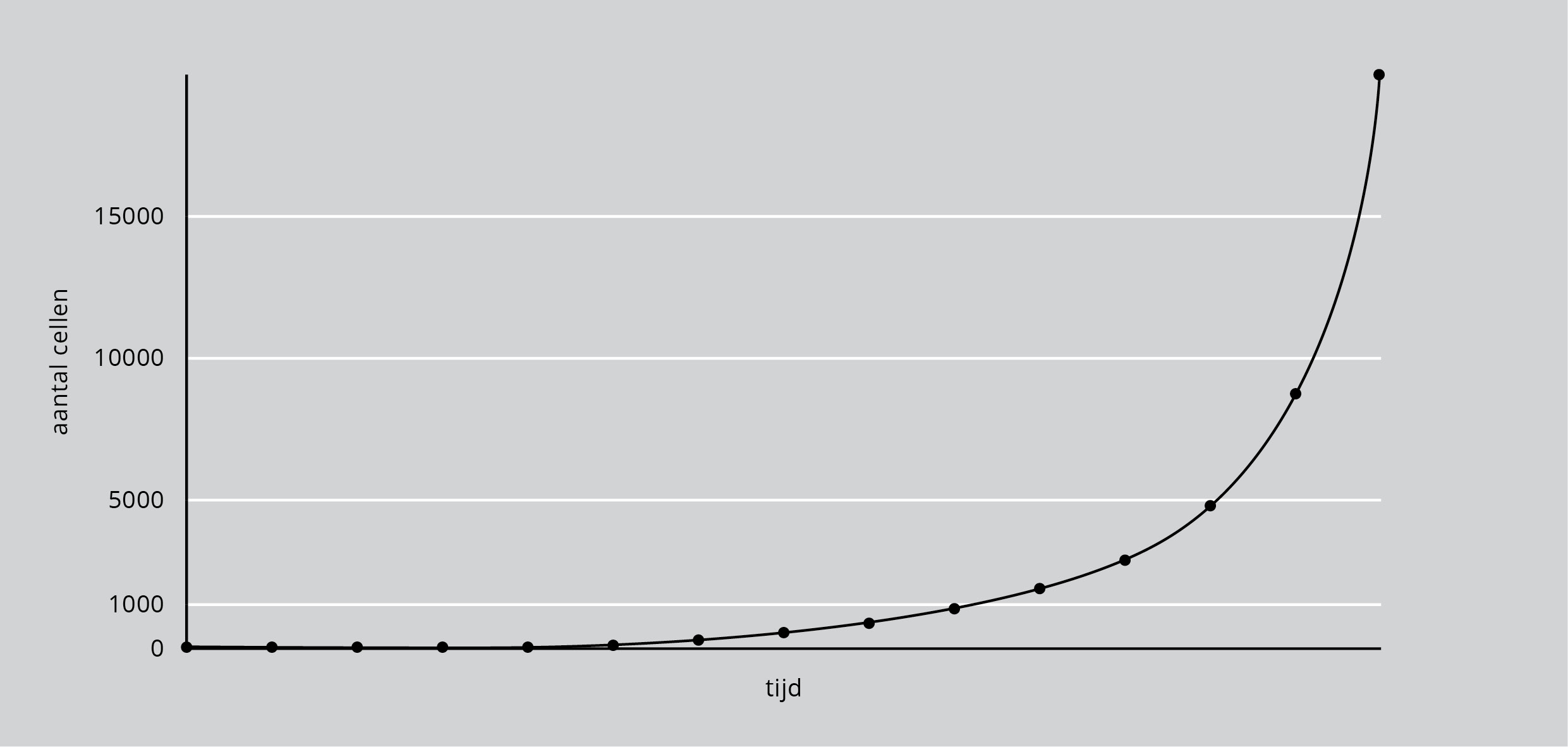
Figuur 1: Exponentiële groei van een populatie waarbij de stippen steeds een vast tijdsverloop weergeven. Eerst is de toename zo langzaam dat het in een grafiek als deze (links onderaan) niet kan worden weergegeven, daarna loopt de lijn bovenaan van het papier af.
Exponentiële groei is een ecologisch èn een economisch sleutelbegrip.

.
Men kan exponentiële groei ook weergeven in procenten. Dat slaat dan op een bepaalde periode. Men heeft het bijvoorbeeld over de bevolkingsgroei van Nederland in 2017, of de economische groei van China in 2017. Een percentage is vaak gering, waardoor men zich niet altijd de werkelijke groei realiseert. Het is in de ecologie daarom de gewoonte uit het jaarlijkse groeicijfer een ander cijfer te berekenen: de verdubbelingstijd van de populatie. Tabel 1 laat enige voorbeelden van verdubbelingstijden zien. Als de populatie met 0,1 procent per jaar toeneemt (dus 1 individu per 1000 individuen erbij), dan zal de populatie elke 700 jaar in aantal verdubbelen. Als het groeitempo 7 procent per jaar bedraagt, verdubbelt de populatie in slechts 10 jaar.
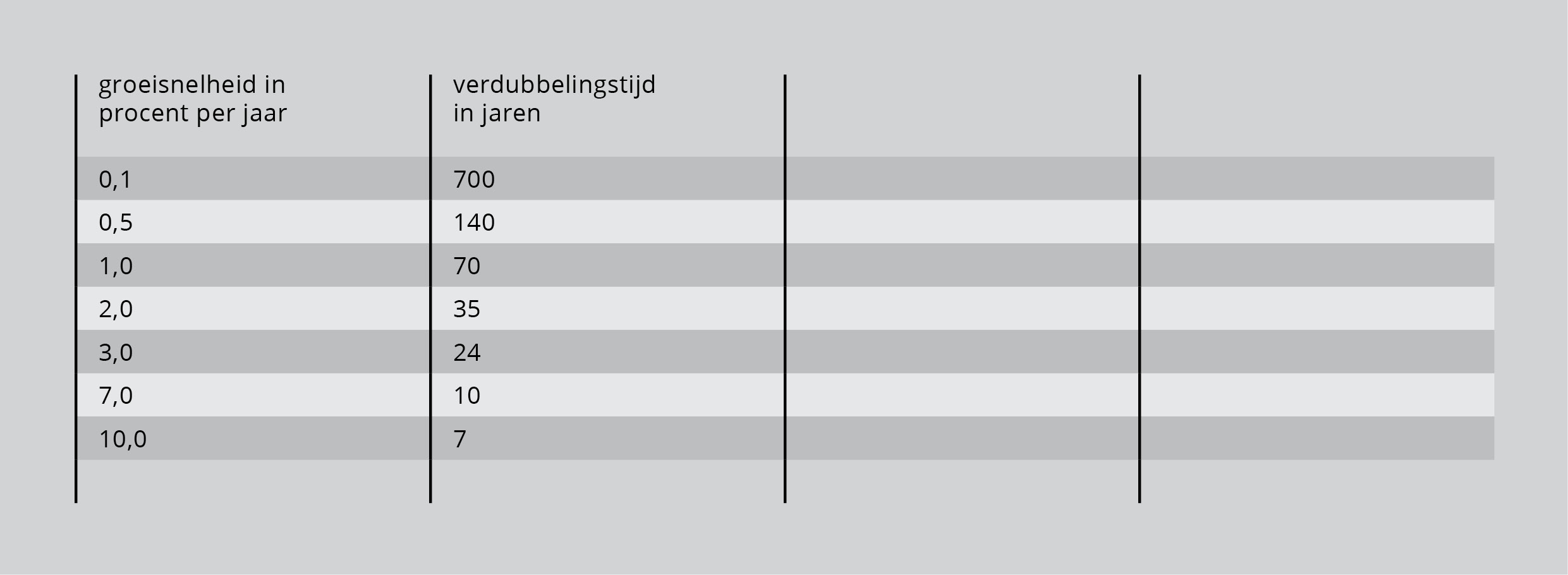
Tabel 1: Enige voorbeelden van verdubbelingstijden.
Vanaf 1990 begon de economie van China explosief te groeien. De op hele procenten afgeronde groeicijfers staan in figuur 2 als vette punten in een lijn. De omvang van de economie is donkergrijs. We zien dat het vanaf 1990 zeven jaar duurt voordat de economie verdubbeld is. De volgende verdubbeling duurt ook (bijna) zeven jaar. De economie is in 2005 dus ruim vier keer zo groot als in 1990. Al in 2011 is de volgende verdubbeling een feit. De economie is nu acht keer zo groot als in 1990. En in 2014 is hij tien keer zo groot als in 1990.
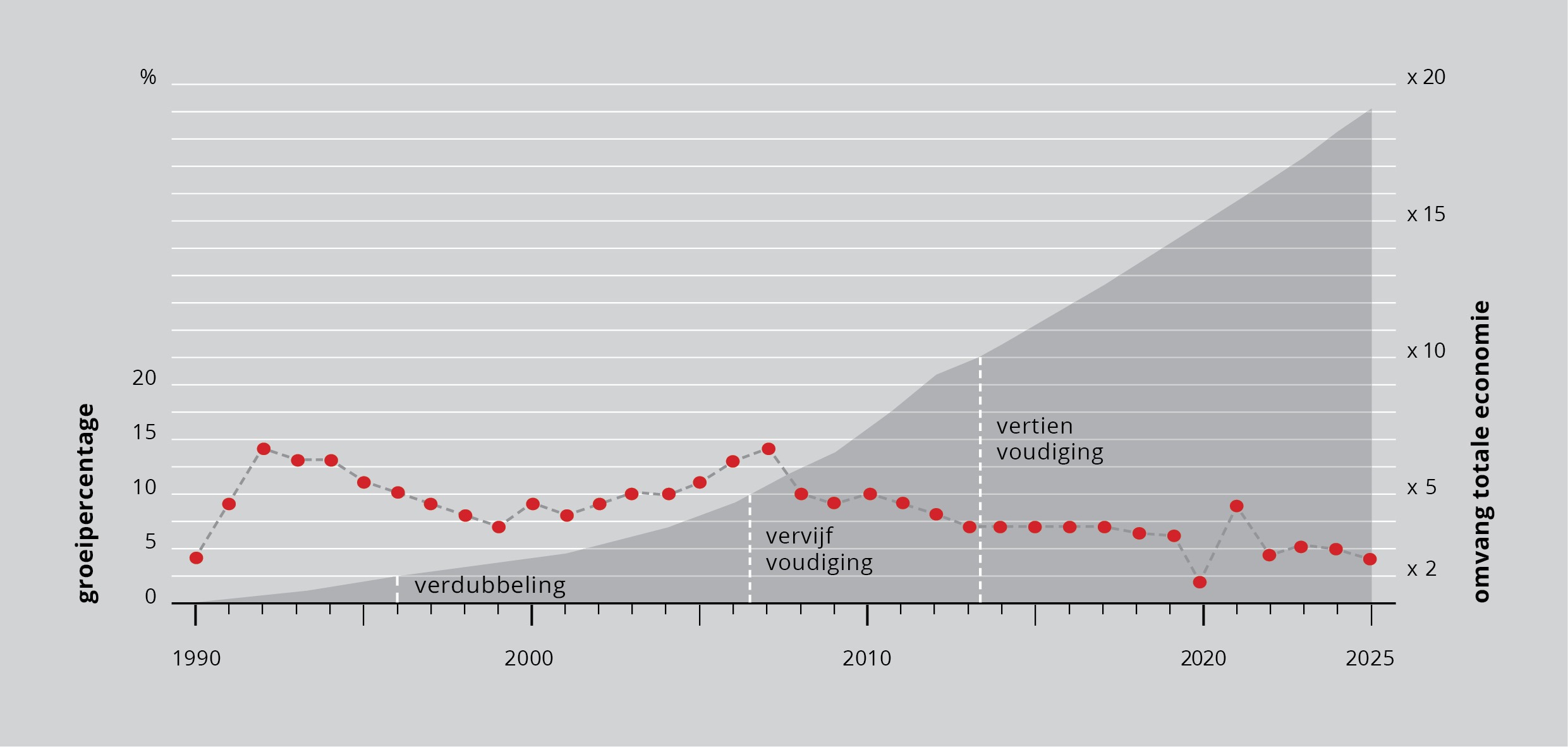
Figuur 2: De economische groei van China.
Als er gezegd wordt “de groei van de Chinese economie is 9%” betekent dat in 1991 dat er bij de 100 eenheden van 1990 9 eenheden bijkomen, maar in 2000 zijn dat er (met eenzelfde groei van 9%) niet 9 maar 23 en in 2011 zijn het er 64. Als er in 2011 wordt gezegd “die 9% economische groei valt tegen” komt er in de praktijk toch zeven keer zoveel economie bij als twintig jaar eerder in 1991. In 1991 was zo’n economische groei geweldig; in 2011 is zeven keer zoveel voor sommigen teleurstellend.
Als de groei 7% zou blijven, is de Chinese economie al in het jaar 2024 twintig keer zo groot als in 1990.
Waarom zie je in de natuur zo weinig exponentiële groei?

.
Het geboortecijfer van veel organismen is buitengewoon hoog. Sommige vissen leggen honderdduizenden eieren; andere organismen, zoals oesters, brengen elke keer dat ze zich voortplanten miljoenen nakomelingen voort. De wereld is niet vol oesters of vissen, dus moeten de sterftecijfers ook erg hoog zijn. Vaak slaagt meer dan 99 procent van de jongen, de eieren of het zaad er niet in om uit te groeien. Van wie dat wel lukt, sterft het gros voordat ze in staat zijn zichzelf voort te planten. Als in een populatie het aantal individuen dat erbij komt, gelijk is aan de sterfte, is er stabiliteit; de bevolkingsgroei is nul. Alleen als het aantal geboorten het aantal sterfgevallen overtreft groeit de populatie.*
Voor de mens wordt daarbij het begrip vruchtbaarheidsgraad gebruikt. Dat is het gemiddelde aantal kinderen per vrouw. Wat nodig is om de bevolkingsomvang in stand houden verschilt per land. In Reunion is een vruchtbaarheidsgraad van 2,04 genoeg om de bevolkingsomvang op peil te houden. In het door AIDS geteisterde Swaziland loopt het op tot 3,35. Het wereldgemiddelde is 2,3. Dus als de vruchtbaarheidsgraad voor alle vrouwen op aarde hoger ligt dan 2,3, groeit de wereldbevolking; bij een lager getal krimpt de wereldbevolking.
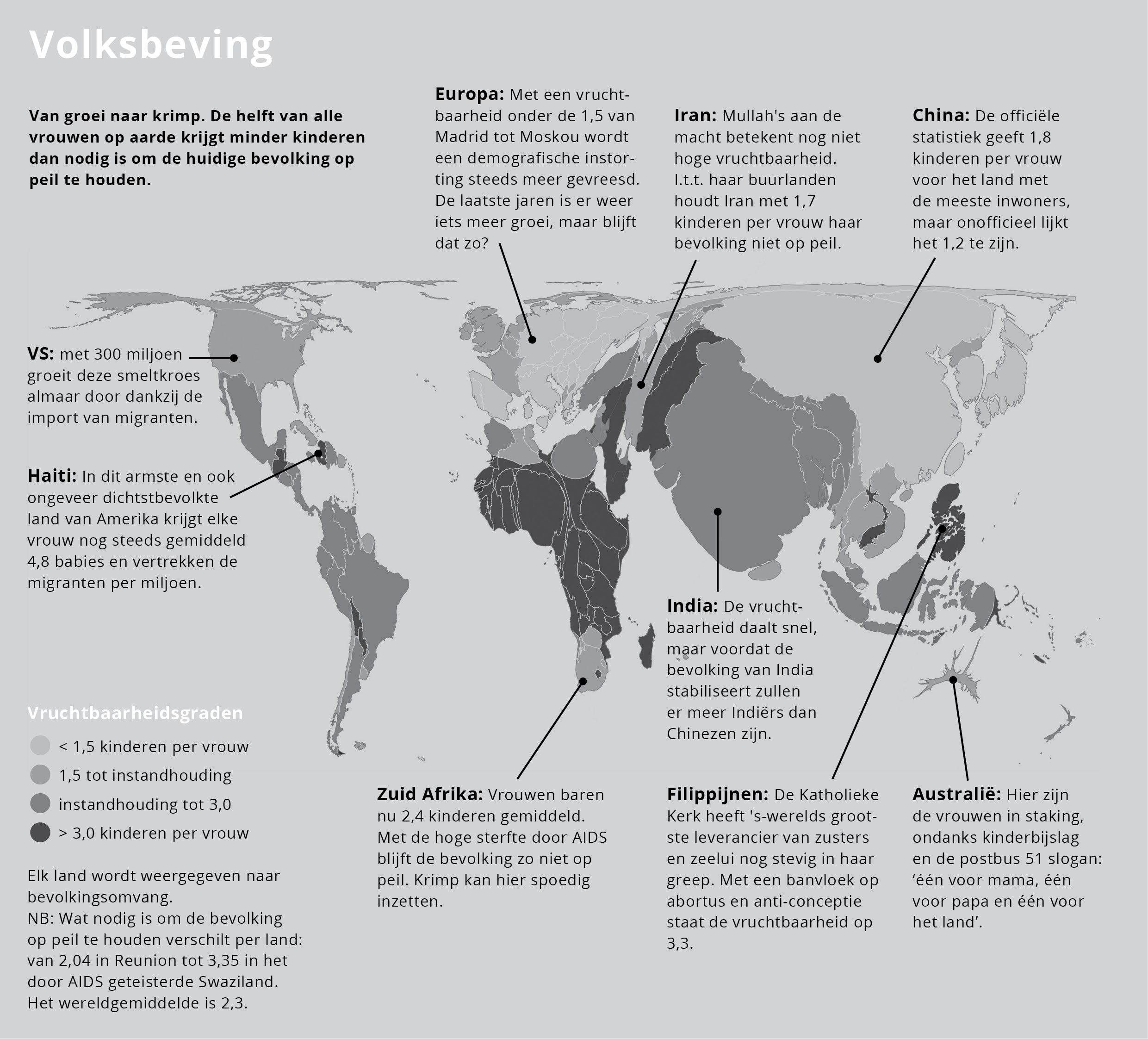
Kaart 1: Vruchtbaarheidsgraden en informatie voor een aantal landen over hun toekomstperspectieven bij de bevolkingsgroei of -krimp.
De trend is dat overal ter wereld vrouwen er nu voor kiezen veel minder kinderen te krijgen dan vroeger. Hierdoor piekte de absolute groei van de wereldbevolking in 1987. Dat jaar kwamen er 87 miljoen mensen bij op aarde; in 2010 was dat 78 miljoen. Die daling gaat door en na 2050 zou de wereldbevolking wel eens kunnen gaan afnemen.
Dat is niet het beeld dat de meeste mensen in hun hoofd hebben. Maar de getallen spreken voor zich: de vruchtbaarheidsgraad van Europa is hooguit 1,5, die van Iran is 1,7, die van China is officieel 1,8 (maar onofficieel misschien slechts 1,2). Ook Zuid Afrika zit op slechts 2,4. Natuurlijk zijn er ook nog ‘haarden van verzet’, meestal waar men fanatiek godsdienstig is en vrouwen weinig te vertellen hebben. Dat is het geval bij de orthodoxe Joden in Israël (verdubbeling in 15 jaar) en de katholieke Filipijnen (vruchtbaarheidsgraad 3,3).
Bij het punt waar elke volgende generatie moeders in aantal kleiner is dan de laatste, wordt een trend van krimp vrijwel onomkeerbaar, net zoals eerst aan de groei niet te ontkomen was. Als elke generatie slechts 1,6 kinderen per vrouw voortbrengt, dan produceren vijf vrouwen slechts vier vrouwen voor de volgende generatie. En zo gaat dat dan door. Meisjes die nooit geboren worden krijgen geen kinderen.
De bevolking van Italië zou (met steeds het huidige vruchtbaarheidscijfer van 1,3 en zonder enige emigratie en immigratie) in een eeuw teruglopen van 58 miljoen (nu) tot slechts 8 miljoen! Niet voor niets wordt in Europa van Madrid tot Moskou een demografische instorting gevreesd.
In de natuur is de kindersterfte zeer hoog.

.
Sinds eeuwen brengt de mens planten en dieren in delen van de wereld, waar ze tot dan niet voorkwamen. Deze introducties van exoten mislukken vaak, maar soms slagen ze. De gevolgen kunnen dan rampzalig zijn voor de inheemse soorten. Vaak lukken introducties op afgelegen eilanden, misschien omdat eilanden gewoonlijk minder soorten bevatten dan het vasteland en daarom meer ontvankelijk zijn. Het ongerepte tropische woud biedt daarentegen krachtig weerstand aan introducties, misschien omdat al grote aantallen soorten aanwezig zijn. De verspreiding van een organisme in een nieuw gebied begint meestal langzaam, maar wint al gauw aan snelheid, ongeveer zoals de groei van het aantal gistcellen in figuur 1 (aflevering 4).
Een sprekend Nederlands voorbeeld is de introductie van de amerikaanse vogelkers (Prunus serotina), die in het begin van de 20ste eeuw is ingevoerd uit Amerika. Deze struik (die ook tot een grote boom kan uitgroeien) werd door de Nederlandse Heidemaatschappij en het Staatsbosbeheer aangeplant in bossen die bestemd waren voor de houtproductie met de bedoeling de bodem te verbeteren. Spoedig bleek deze soort zich echter massaal uit te zaaien en de inlandse Prunussoorten ernstig te beconcurreren. De amerikaanse vogelkers kende hier geen natuurlijke consumenten, in tegenstelling tot de inheemse soorten. Bovendien bleek het strooisel van amerikaanse vogelkers de groei van andere soorten te remmen. Eenmaal geïntroduceerd, lukte het niet er weer af te komen. De ondergroei in veel bossen van voornamelijk amerikaanse vogelkers kreeg daarom de veelzeggende naam ‘bospest’.
Tijdens een epidemie vertonen populaties van ziekte-organismen een spectaculaire groei. Zo is het bij griep. Griep wordt veroorzaakt door een virus. De ziekte slaat steeds opnieuw toe bij miljoenen mensen in grote delen van de wereld. De ziekte verdwijnt even snel als hij verschijnt, maar keert na verloop van misschien een jaar weer terug in een nieuwe vorm, waartegen de mens in het begin niet immuun is. Tegenwoordig zijn vaccins snel gemaakt, maar tijdens de eerste wereldoorlog zijn ongeveer honderd miljoen mensen aan griep gestorven. Dit geeft een idee van de explosieve aard van een epidemische ziekte als deze eenmaal vaste voet gevonden heeft.
Explosieve groei vinden we in allerlei gedaantes.

.
De aanplant van de grove den werd op onze heidevelden en stuifzanden ooit gecombineerd met de introductie van de vogelkers. Vervolgens vond men die hardnekkige vogelkers in de weg staan en noemde men het een plaag of pest.
Vandaag de dag kijken ecologen heel anders tegen deze ‘bospest’ aan. In het boek Amerikaanse vogelkers* stellen de auteurs dat de kijk op de vogelkers als een pest eerder is ingegeven als rechtvaardiging voor de keuze om hem uit te roeien dan dat het een ecologisch inzicht is.
“Als je bossen met grove dennen aanplant, moet je niet verbaasd zijn dat daaronder alleen vogelkers wil groeien. De grove den laat relatief veel licht door en dat heeft de vogelkers nodig. Het heeft weinig zin om de soort krampachtig te bestrijden. Beter is het bossen rijker te maken, met een grotere soortenvariëteit. Als je eik, beuk, esdoorn, lijsterbes en hazelaar plant, dan wordt de concurrentie voor de vogelkers zo groot, dat hij vanzelf minder dominant wordt.”
Ondanks miljoenen zaden van de vogelkers zie je dan maar een enkel stammetje opschieten. “De schaduw van de eiken is al voldoende. Esdoorns en hazelaars verspreiden hun zaad in de wijdere omgeving en hun nakomelingen komen op tussen de vogelkers. Plant je nog meer soorten, dan krijg je vanzelf een weerbaar bos met een gelaagdheid die de vogelkers automatisch tot de orde roept.”
De auteurs van het boek constateren dat “we in Nederland dertig jaar lang de vogelkers hebben bestreden zonder er goed over na te denken en dat het blijkbaar moeilijk is om het vijandbeeld van bospest los te laten.”
We leren hieruit dat we een open blik op de werkelijkheid moeten houden en onze inzichten moeten blijven toetsen.
Ook in de ecologie moeten we oppassen voor tunnelvisie.

.
Fazanten komen oorspronkelijk uit Azië. Ze zijn hier zo bekend dat de meeste mensen denken dat ze er altijd zijn geweest, maar de mens heeft verscheidene soorten fazanten over de hele wereld verspreid. Ze dienen voor de jacht en als voedsel. De meest voorkomende soort is de gewone bosfazant (Phasianus colchicus). In de gematigde streken van het noordelijk halfrond broedt de bosfazant in het voorjaar met een broedsel van 8-15 eieren. Het vrouwtje voedt en verzorgt de jongen, die na ongeveer dertien dagen kunnen vliegen.
Een enkele keer is het mogelijk gebleken om de groei van een populatie te volgen vanaf de oorspronkelijke introductie. In 1937 werden twee mannetjes en zes vrouwtjes ingevoerd op het eiland Protection voor de kust van Washington in het noordwesten van de Verenigde Staten. Er waren daarvoor geen fazanten op het eiland, maar het milieu moet er gunstig voor hen geweest zijn, want de populatie groeide in zes jaar tijd van 8 tot 1898 exemplaren. De populatie werd elk voorjaar en elke herfst geteld en de getelde aantallen zijn weergegeven in figuur 3.
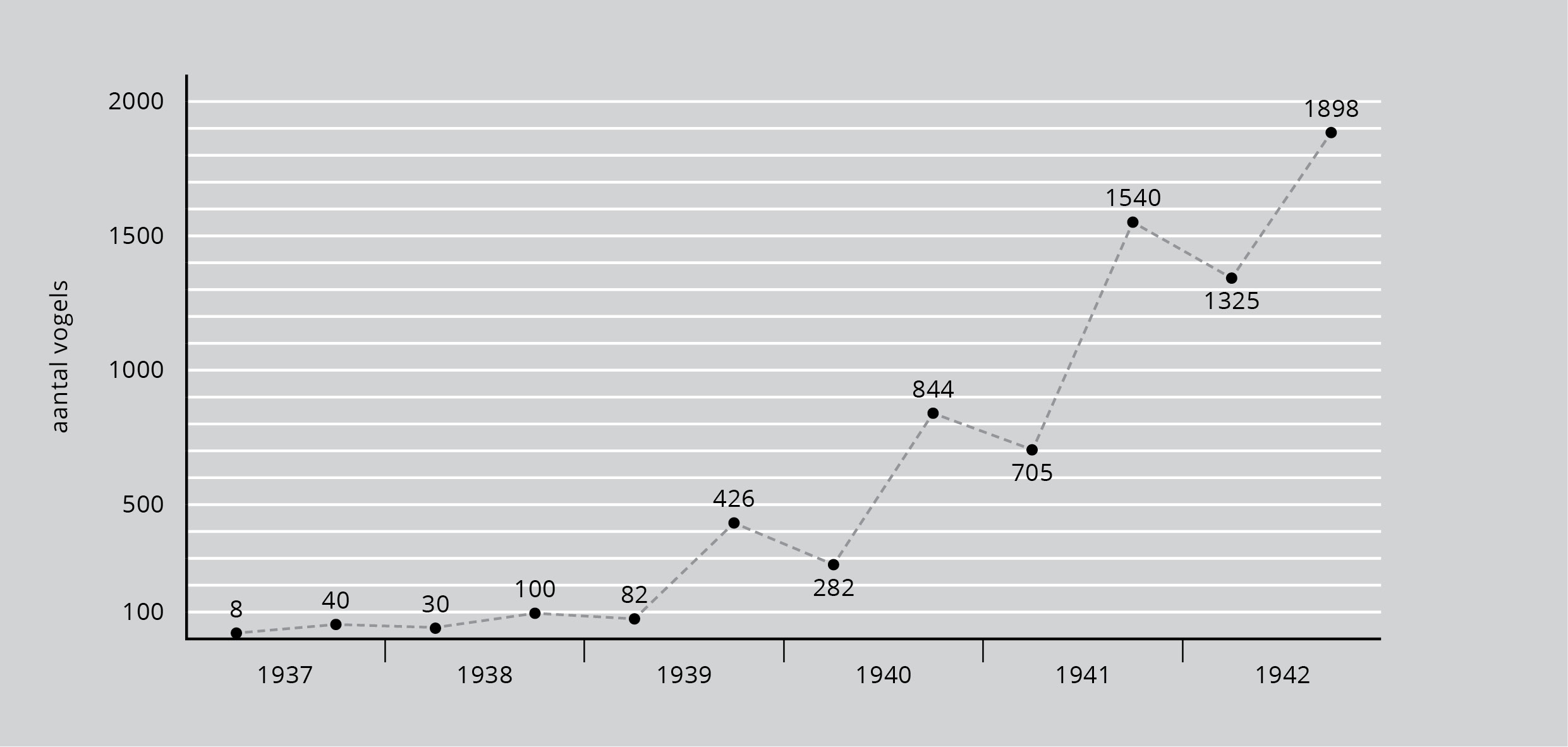
Figuur 3: Toename van het aantal fazanten op Protection Island, Washington. Elke lente en elke herfst werd er een telling uitgevoerd. (Bron: D. Lack, 1954)
Aanvankelijk groeide de populatie langzaam, maar de snelheid nam al gauw toe volgens een ons nu vertrouwd patroon. Zoals figuur 3 laat zien waren er met de lentetelling steeds duidelijk minder fazanten, zonder twijfel als gevolg van de sterfte tijdens de voorafgaande winter. Hierdoor nam de populatie trapsgewijs toe. Na 1942 beschikken we niet meer over gegevens, maar het groeitempo begint al tekenen van verflauwing te vertonen. Op langere termijn blijft een populatie niet doorgroeien. De groei van de fazantenpopulatie op Protection Eiland is dezelfde als die van de gistcellen uit figuur 1 in aflevering 4).
In zes jaar tijd van 8 naar 1898.

.
Wanneer mensen proberen een plaag of onkruid te bedwingen zien we steeds hetzelfde patroon. Als bijvoorbeeld een tuin wordt overspoeld met een of ander hardnekkig onkruid, zoals zuring (Rumex), kan een poging worden gedaan om het uit te roeien. Gebruikt men daarvoor een bestrijdingsmiddel, dan zal dat de eerste keer praktisch alle planten doden en de tuinier zal het gevoel hebben dat het bij de eerste poging gelukt is. Maar de kans bestaat dat enkele planten na een tijdje weer tevoorschijn komen en wordt er verder niets tegen gedaan, dan zal het onkruid zich weer over de tuin verspreiden en is er al gauw weer evenveel als vroeger.
Je kunt proberen het bestrijdingsmiddel bij herhaling toe te passen, in de hoop dat het onkruid tenslotte geheel zal verdwijnen. Dit kan heel lang duren, omdat het onkruid een opmerkelijk en ergerlijk vermogen toont om te overleven en de kop weer op te steken. De exponentiële afname van het onkruid in de tuin wordt gevolgd door een zeer moeizame strijd tegen de laatste resten. Want een paar overlevende organismen kunnen de populatie weer opbouwen tot het oorspronkelijke aantal.
Exponentiële groei van een populatie kan dus leiden tot onverwacht grote aantallen in korte tijd. Exponentiële afname kan bij oppervlakkig onderzoek de schijn wekken dat een populatie verdwenen is.
Je kunt de bestrijding ook aan de natuur overlaten. We kijken wat er gebeurt aan de hand van lijsters en slakken. Lijstersoorten als de merel en de zanglijster zijn ervaren slakkenvangers, vooral van de grotere soorten slakken die in tuinen en heggen leven, zoals de tuinslak (Helix aspersa) en van twee plattelandssoorten (Cepaea nemoralis en Cepaea hortensis). De lijsters eten slakken voornamelijk bij droog weer, wanneer wormen, hun favoriete voedsel, moeilijk te vinden zijn.
Als de vogel een slak vindt, neemt hij deze mee naar een steen, waarop hij het huisje verbrijzelt om bij het zachte, eetbare lichaam ervan te komen.
In de tuin van ons experiment wordt steeds weer dezélfde steen gebruikt. Vogelkenners noemen zulke stenen soms ‘slakkenaambeelden’. Door het verzamelen van de stukken van de lege slakkenhuisjes rond de aambeelden, is het mogelijk uit te vinden hoeveel slakken er zijn opgegeten.
Over het experiment lees je in de volgende aflevering. Daar blijkt dat lijsters meer slakken eten als er veel van zijn. Met andere woorden het consumptietempo van een bepaalde voedselbron hangt samen met de aanwezige hoeveelheid ervan. (In het experiment was de tijdsduur kort genoeg om te zorgen dat het aantal lijsters gelijk bleef en dat het aantal slakken alleen door de lijsters werd beïnvloed.)
Exponentiële afname eindigt meestal ver boven nul.

.
In mei 1972 werden op een plek in Zweden, waar geen lijsters voorkomen, 724 exemplaren van de slakkensoort Cepaea nemoralis verzameld. Deze werden vervolgens naar een tuin in de Engelse Midlands overgebracht, waar verscheidene lijsters huisden. De slakken werden op 23 mei losgelaten en begonnen zich onmiddellijk in de plantengroei te verspreiden. Omdat het slakken waren gingen ze niet erg ver.
Voor die tijd waren in de tuin slechts enkele exemplaren van de soort Helix aspersa bekend. Waarschijnlijk hadden de lijsters nog nooit eerder slakken van de Cepaea-soort gevonden. De lijsters ontdekten de slakken pas na vier dagen, op 27 mei. Ze aten er die dag 90, dat is 12,4% van het totaal. Vanaf dat moment vonden de vogels elke dag slakken en binnen zestien dagen hadden ze er 630 te pakken, bijna 87% van het totaal.
Tabel 3 laat onder meer het aantal slakken zien dat elke dag werd gegeten. In het begin van het experiment, dus toen er nog veel slakken waren, vingen de lijsters de meeste slakken. In de eerste vijf dagen aten ze bijna 70% van het losgelaten aantal. De laatste kolom laat zien welke gedeelte van de nog overgebleven slakken werd gevangen. De eerste week vingen de lijsters elke dag minstens 12% van het totale aantal slakken in de tuin. Daarna daalde het aantal gevangen slakken abrupt, totdat tegen het eind van het experiment het aantal slakken dat gevangen werd minimaal was. Toch waren er toen nog 90 slakken over.
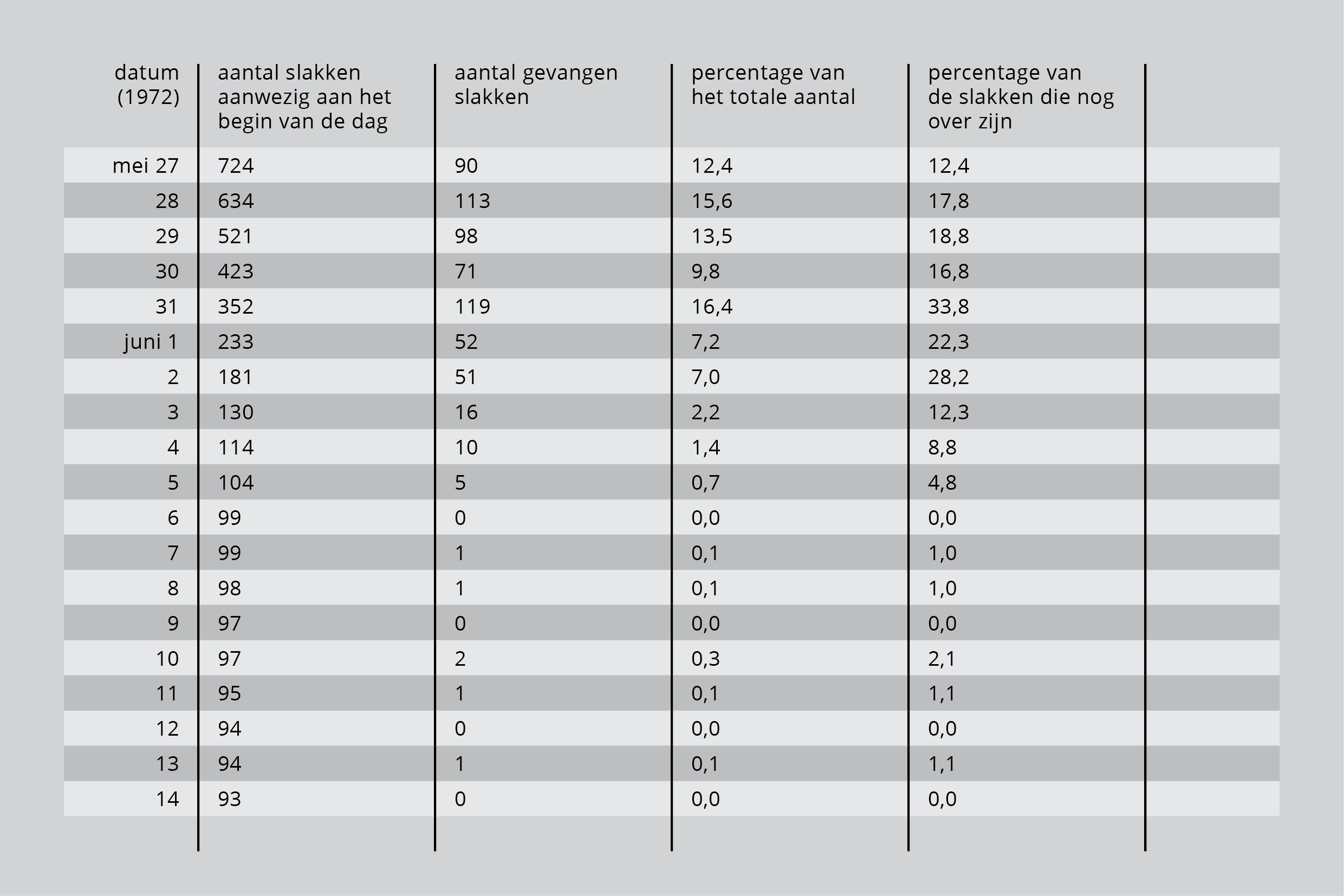
Tabel 2: Slakken die door de lijsters op achtereenvolgende dagen gevangen worden.
Lijsters zijn dus het meest efficiënt zolang er slakken in overvloed zijn. Opmerkelijk is dat wanneer de slakken schaarser worden, de lijsters er van de overblijvers naar verhóuding minder vinden! We moeten in gedachten houden dat naarmate er tijd verliep de slakken zich misschien beter gingen verbergen, of dat de slakken die zich direct al het beste verborgen degene waren die bleven leven.
Tussen 12 juni en 15 november werden nog 13 exemplaren gevangen en het is waarschijnlijk dat enkele slakken in deze periode aan andere oorzaken dood gingen. Maar een jaar later, in juni 1973, waren er nog altijd een paar levende slakken in de tuin. De lijsters waren allang opgehouden naar hen te zoeken, en waren weer overgegaan op het eten van wormen en insecten.
Het experiment laat zien dat de consumptie het omgekeerde verloop vertoont van de exponentiële groeicurve. Eerst is het verbruik hoog, maar naarmate het voedsel wordt opgegeten, neemt het consumptietempo af.
Wordt een voedselbron schaars dan is er minder kans op dat hij wordt geëxploiteerd.
.
Als we het over levensbehoeften hebben, denken we voornamelijk aan de beschikbaarheid van energie en voedingsstoffen. Groene planten vangen stralingsenergie op van de zon en zetten deze om in opgeslagen chemische energie. Dit gebeurt met het proces van de fotosynthese.
Alle andere organismen zijn voor hun voedsel direct of indirect afhankelijk van de groene planten. Men zou daarom denken dat dierpopulaties tenslotte beperkt worden door het vermogen van de planten om zonne-energie om te zetten in voedsel, maar dat is te simpel gesteld.
Er zijn veel manieren waarop de individuele leden van alle planten- en dierpopulaties op elkaar inwerken. Zo kunnen ze met elkaar strijden om dezelfde energie- en voedselbronnen, of om dezelfde ruimte. Dit kan zover gaan dat de overlevingskansen van één individu afhangen van het aantal mededingers dat tegelijkertijd aanwezig is. Hoe groot een populatie kan worden hangt af van het vermogen van haar leden om aan roofdieren en parasieten te ontsnappen, hoeveel weerstand ze bezitten tegen ziekten en hoe goed ze zich weten te verbergen.
Er zijn dus veel factoren van invloed op de omvang van een populatie. Lang niet allemaal hebben ze direct te maken met de voorziening van energie en voedsel. Daarom gebruikt men vaak de term ‘draagkracht’ om mee aan te geven wat het vermogen van een milieu is om een populatie te onderhouden. Bij deze draagkracht van het milieu moeten we een bepaalde populatie bekijken en alles in beschouwing nemen wat van invloed is op de omvang van die populatie. Meestal blijken er voor die populatie een paar factoren werkelijk van belang te zijn. Men noemt ze sleutelfactoren. Als die veranderen heeft dat direct invloed op de omvang van die populatie. Als andere factoren veranderen, wijzigt de omvang van de populatie veel minder.
Een voorbeeld vormt de algengroei in de sloten. Daarbij is de hoeveelheid fosfaat in het water de doorslaggevende factor. Van nature zit er maar weinig fosfaat in het water en hierdoor wordt de algengroei geremd. Als uit mest fosfaat in het water komt, wordt dit tekort opgeheven. Er ontstaat een nieuwe situatie, waarin andere factoren van doorslaggevende betekenis zijn geworden.
De ene doorslaggevende factor kan opgevolgd worden door een andere.

.
Wanneer een populatie voor het eerst ergens ontstaat, zal deze exponentieel groeien (denk aan de fazanten), maar onder normale omstandigheden wordt al snel de draagkracht van het milieu bereikt. Dat wil zeggen, bepaalde factoren zullen een sterke belemmering gaan vormen voor verdere groei van de populatie. Als de draagkracht wordt bereikt, zal de populatiegroei afnemen en tenslotte zullen sterftecijfer en geboortecijfer gelijk zijn. Er vindt dan geen verdere groei plaats en de populatie is op een bepaald niveau ‘gereguleerd’.
De grafiekjes van figuur 4 laten verschillende mogelijke regulatieprocessen zien. In de praktijk zijn geboorte en sterfte van de meeste populaties al lang in evenwicht, met alleen kleine schommelingen in de omvang van de populatie. In elk van de drie grafiekjes wordt de populatie weergegeven door het aantal exemplaren. Het is moeilijk de ‘eenheid van draagkracht’ van het milieu te geven. De draagkracht is eenvoudig een maat voor het vermogen van het milieu om een populatie van een of andere soort gedurende een bepaalde periode te onderhouden. Maar men kan de draagkracht in de praktijk meestal gelijk stellen aan de beschikbaarheid van voedsel.
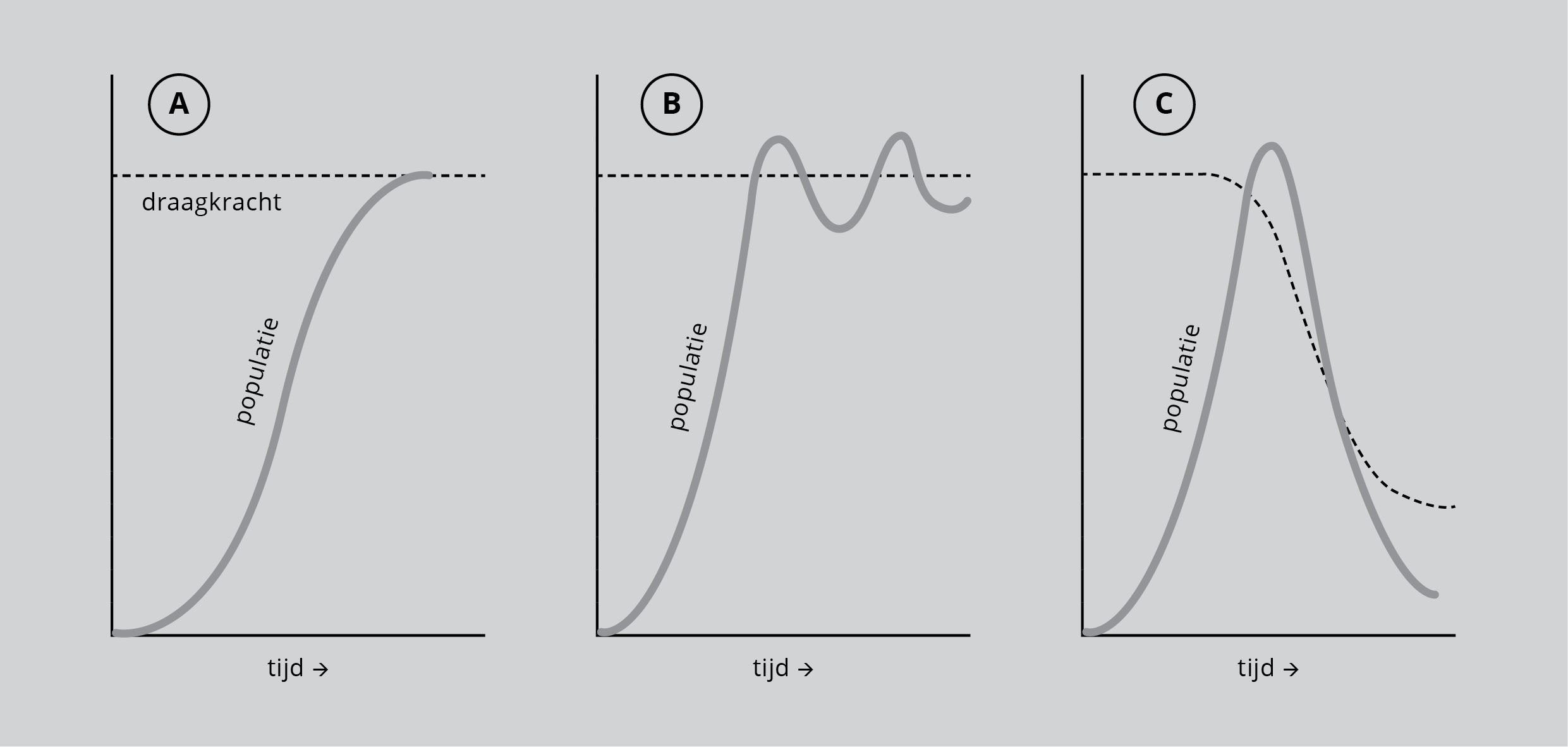
Figuur 4: A Groei van een populatie tot een stabiel niveau als gevolg van de beperking die de draagkracht van het milieu uitoefent. B: Groei van een populatie tot een stabiel niveau dat zich beweegt rondom de draagkracht van het milieu. C: Groei en afname van een populatie doordat de populatie afbreuk doet aan de draagkracht van het milieu.
In figuur 4A en 4B groeit de populatie steeds eerst snel, maar als de draagkracht wordt genaderd neemt het groeitempo af, om daarna te stoppen of te schommelen. De populatie is dan in evenwicht met de draagkracht van het milieu. Dezelfde exponentiële groei komt voor in figuur 4C, maar deze keer wordt de draagkracht aangetast door de druk die de populatie uitoefent en is er daarna een afname van zowel de draagkracht als van de populatie. Men kan hier ook zeggen dat de populatie zijn eigen grondstoffen uitput. Het evenwicht kan zich dan op verschillende manieren verder ontwikkelen. Zo zou de populatie door haar eigen druk te gronde kunnen gaan. De draagkracht wordt dan bevrijd van de druk die de populatie uitoefent en kan weer omhoog gaan tot haar oude niveau. Ook zouden zowel de populatie als de draagkracht door kunnen gaan op een lager niveau dan eerst.
Als we de omvang van een populatie en de draagkracht van het milieu over een langere periode meten, zien we bijna altijd dat het patroon dan min of meer in evenwicht is. Zo is het meestal, ondanks het vermogen van populaties om exponentieel te groeien.
Onze economische groei lijkt de draagkracht van de aarde bereikt te hebben of al te overschrijden.

.
In de lente als de bomen weer groen worden, gaan vele soorten zangvogels meer zingen. Onderling verschilt de zang van de soorten aanzienlijk. De zang is dan ook vooral bedoeld voor soortgenoten. Eerst lokt het mannetje met zijn zang een wijfje. Later als de vogels gaan broeden, is het bij de zangvogels vooral het mannetje dat met zijn zang een gedeelte van het bos afbakent: dit stuk is bezet. Er is een bepaald stuk bos als voedselgebied nodig, want in het bos is de voedselvoorraad niet onbeperkt. Er is hier dus sprake van concurrentie tussen vogels van dezelfde soort om een stuk van het bos. Zo’n eigen stuk bos heet een territorium.
Eén van de zangvogels in het bos is de vink. De vink heeft zijn territorium liever in een gemengd bos dan in een naaldbos, vanwege het voedsel dat er voorkomt en de betere nestgelegenheid die het de vink biedt. Dus wordt eerst het gemengde bos bevolkt. Als een bepaald aantal territoria in het gemengde bos bezet is, wordt er door de populatie overgestapt op het minder gunstige naaldbos. Men kan zeggen dat de draagkracht van het gemengde bos wordt benaderd. De draagkracht van het naaldbos is voor vinken geringer dan de draagkracht van het gemengde bos. De territoria in het naaldbos zijn groter, de grenzen ertussen minder duidelijk. In de aantallen vinken in het gemengde bos verandert jaarlijks weinig, want alle mogelijke territoria worden steeds gevuld. In het naaldbos schommelt het aantal vinken meer, want het minst gunstige gebied zal pas bij hoge ruimte- en voedselnood bezet worden. Dit komt tot uitdrukking in tabel 3 en figuur 5.
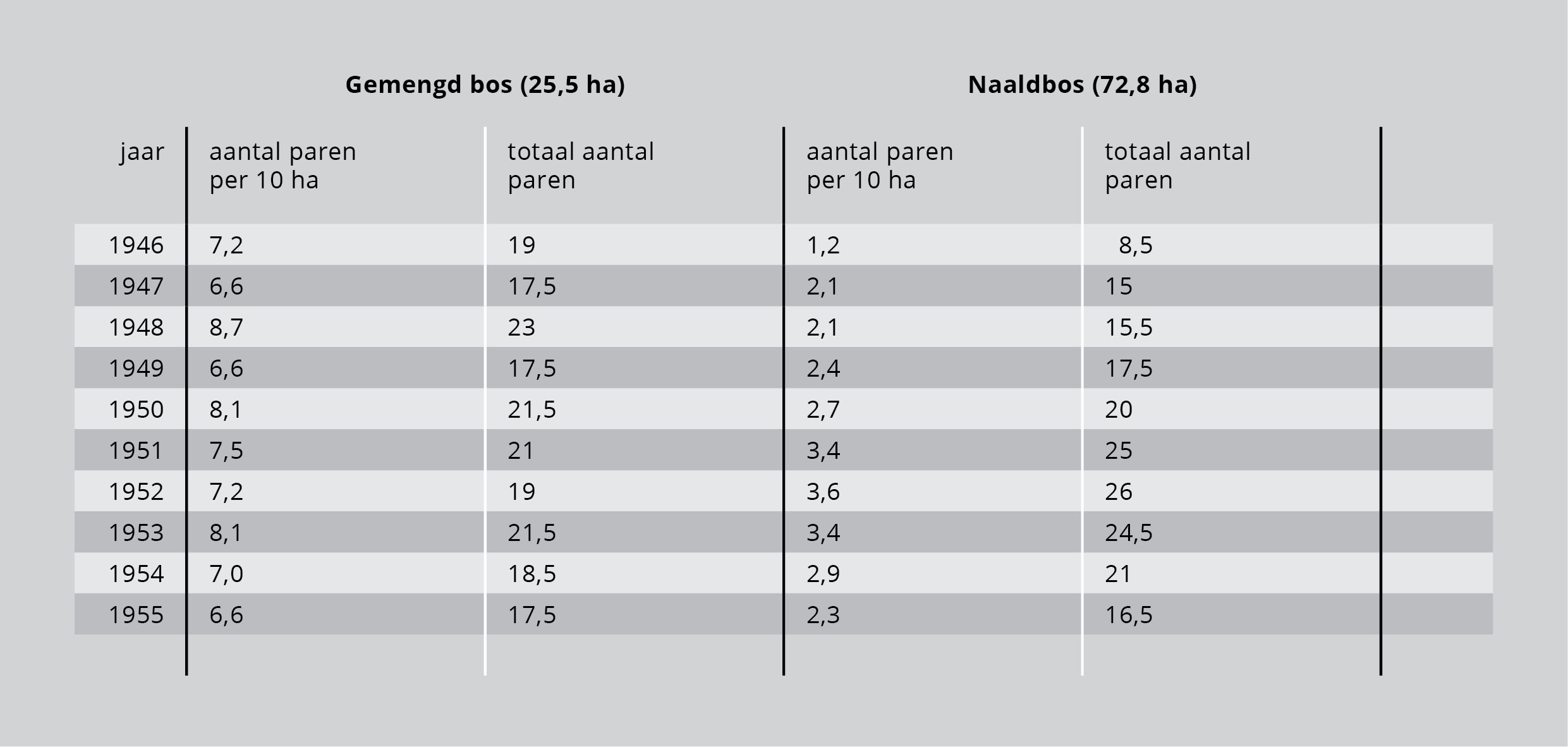
Tabel 3: De fluctuerende populatiedichtheid van vinkenmannetjes in respectievelijk loofbos en naaldbos gedurende tien opeenvolgende jaren in het Hulsthorster Zand (bron: P. Glas, 1960).
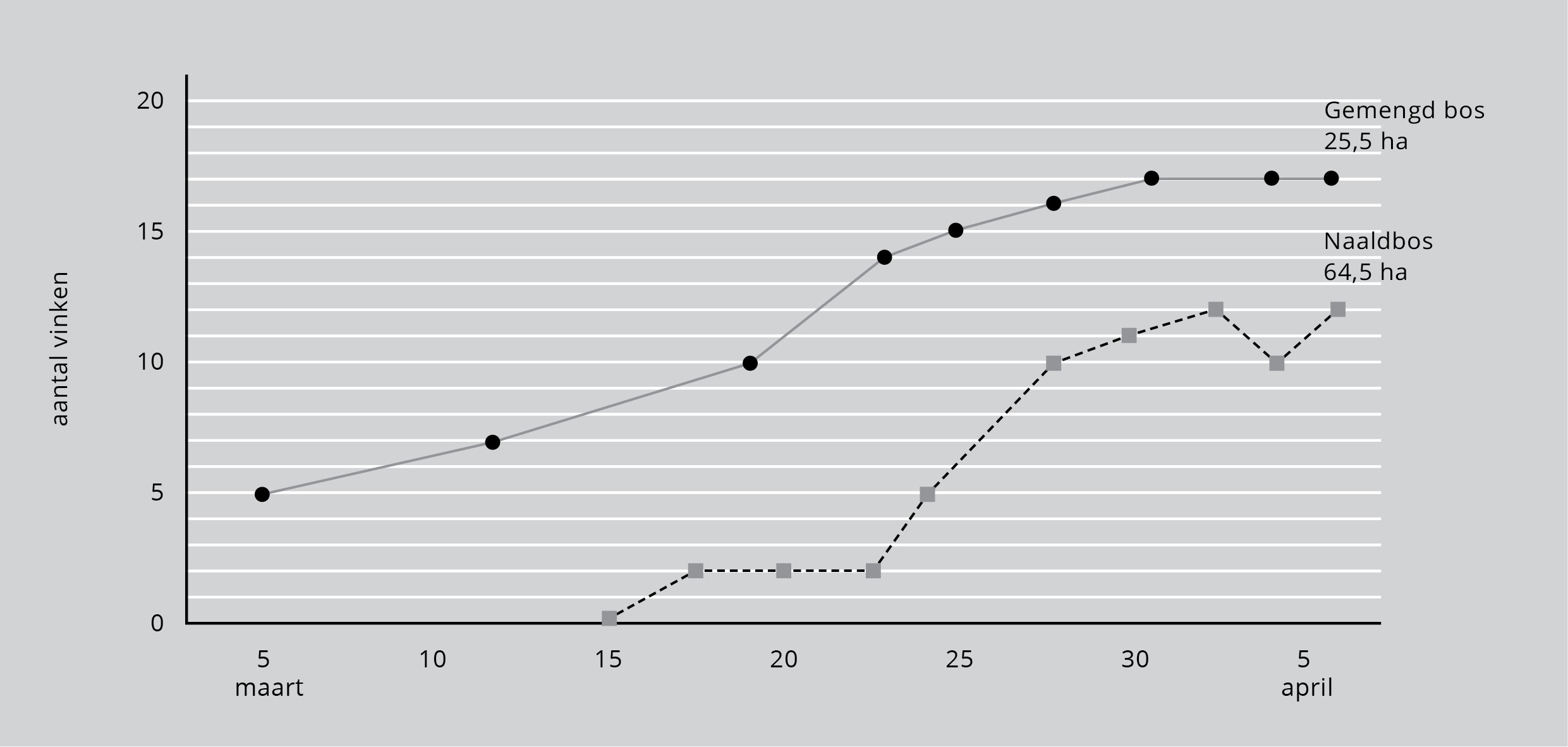
Figuur 5: Het verloop van de bezetting van een gemengd bos en een naaldbos door mannetjesvinken (bron: P. Glas, 1960).
Draagkracht is geen neutraal verschijnsel maar keiharde realiteit.

.
In de natuur geldt steeds dat individuen met elkaar concurreren omdat de draagkracht van het milieu beperkt is. Dit concurreren hoeft niet noodzakelijk te betekenen dat individuen openlijk met elkaar in conflict zijn. Men ziet onder zoogdieren en vogels van één soort zelden gevechten op leven en dood. Wel zie je vaak dreigend gedrag. Denk eens aan de roodborstjes in het vogelhuisje in de winter. Zij verjagen vaak een ander van het voedsel. Daarbij reageert de ene vogel prompt op het dreigende gedrag van de ander en gaat opzij. Het ligt voor de hand dat een dier dat steeds bij het voedsel wordt weggehouden, gebrek zal lijden en dat dit in tijden van voedselschaarste de dood kan betekenen. Evenzo zal de sterkste en grootste plant de zwakste verdringen als er maar plaats is voor een.
Onder concurrentie verstaan we dus een toestand waarbij individuen van één soort, grondstoffen consumeren die maar beperkt beschikbaar zijn. De bekwaamste zullen overleven; de rest zal wegkwijnen.
De concurrentie is groter naarmate de populatie dichter is. Want hoe meer individuen, des te groter de kans dat de draagkracht van het milieu onder druk staat. De overlevingskans hangt dus samen met de dichtheid van de populatie. Ecologen zeggen daarom vaak dat het sterftecijfer dichtheidsafhankelijk is. De beschikbaarheid van voedsel zal vermoedelijk de overlevingskans dichtheidsafhankelijk beïnvloeden.
Andere factoren werken onafhankelijk van de dichtheid. Bijvoorbeeld ongebruikelijke weersomstandigheden zoals droogte of overstromingen zullen alle leden van de populatie raken. Vogelpopulaties in bosgebieden lijken vooral onder de druk van de dichtheidsafhankelijke factoren te staan, terwijl voor insecten in droge gebieden geldt dat de sterfteoorzaak meer aan dichtheidsonafhankelijke factoren valt toe te schrijven.
Er is altijd concurrentie.

.
Heeft het aantal geboorten gevolgen voor de stabiliteit van populaties? Planten en dieren die in het gematigd klimaat leven, planten zich meestal op vaste tijden van het jaar voort. We kennen allemaal de jaarlijkse cyclus van bloeien en vrucht dragen, van het broeden van vogels in de lente en van het tevoorschijn komen van insecten in de zomer.
In de tropen is het iets ingewikkelder. Daar komt voortplanting bij veel soorten gedurende het hele jaar voor, maar bij bijna alle soorten vinden we ook daar seizoenpieken in de voortplantingsactiviteiten.
Voortplanting in vaste seizoenen doet in een populatie plotseling grote aantallen jonge planten of dieren verschijnen. De meeste lukt het slechts korte tijd te blijven leven. Slechts een paar van de plantenzaadjes groeien uit tot volwassen exemplaren die zichzelf kunnen voortplanten. Ook de meeste eieren en jongen brengen het niet tot geslachtsrijpe volwassenen. Het sterftecijfer van de jongen is buitengewoon hoog, vaak meer dan 99%.
Neem een vrouwelijke vlinder die 200 eitjes legt. Als de vlinderpopulatie stabiel moet blijven, zal tot de volgende generatie gemiddeld slechts één volwassen mannetje en vrouwtje mogen overleven. De overblijvende 198 ‘moeten’ sterven als eitjes, rupsen of als pop. Sterfte komt in alle ontwikkelingsstadia voor, maar onderzoek naar de doodsoorzaken laat zien dat de jongen het ergst te lijden hebben.
Als een vlinder, vogel, boom of wat voor organisme dan ook eenmaal volwassen is geworden en in staat is zich voort te planten, zijn de kansen om te blijven leven sterk verbeterd. (Voor sommige organismen zoals virussen en gistcellen heeft het begrip ‘volwassen zijn’ weinig betekenis. Er bestáát geen duidelijk volwassen stadium.)
Laten we dit eens bekijken bij de koolmees.
De meeste nieuwe planten en dieren sterven jong.

.
De koolmees broedt meestal in boomholtes, maar met nestkastjes is het een stuk gemakkelijker om gegevens te verzamelen over het aantal eieren dat wordt gelegd en het aantal jongen dat daar uit komt.
De koolmees legt 10-12 eieren. De meeste ervan komen uit en wel op dezelfde dag. De jongen worden door de ouders gedurende iets meer dan twee weken gevoed met de rupsen die meestal rijkelijk voorkomen op de bladeren van bomen zoals de eik en de iep.
In de vroege zomer vliegen de jonge vogels uit. Ze voeden zich met verschillende insecten. In de meeste jaren ontstaat er tegen juli en augustus een voedseltekort door gebrek aan insecten. Als de jongen uitvliegen is er een soort van bevolkingsexplosie in het bos. Elk paar heeft 10-12 jongen voortgebracht, zodat in een stuk bos van 20 broedende paren de totale populatie ineens 240-280 mezen kan bedragen.
Toch is bij het begin van de winter de populatie weer terug op ongeveer 40 vogels. Weinig daarvan zijn van het broedsel van dat jaar. Anders gezegd: het zijn eerder de ouders die overleven dan hun jongen. Er moeten veel mezen gestorven zijn. De doodsoorzaken zijn niet gemakkelijk te bepalen, omdat in de zomer de bossen zo vol groen staan en de vogels zo stil zijn dat ze nauwelijks opvallen. Hoe dan ook, het is duidelijk dat de explosie niet leidt tot een grote populatie in de winter en dat voor het volgende broedseizoen de populatie weer is gezakt tot het oude niveau. Er zijn natuurlijk van jaar tot jaar schommelingen, maar het aantal wordt nooit zo groot als je zou verwachten met het aantal jongen dat uitvliegt.
Figuur 6 laat dit zien aan de hand van een populatie koolmezen in een klein bos bij Oxford (Engeland). De vogels werden in de winter geteld, als ze gemakkelijk te zien waren en opnieuw als ze in het voorjaar in de nestkastjes broedden. Het aantal jongen was zo bekend. Dit maakte een schatting voor de vroege zomer mogelijk. Een werkelijke telling kon pas weer in de herfst plaatsvinden, wanneer de bladeren waren gevallen.
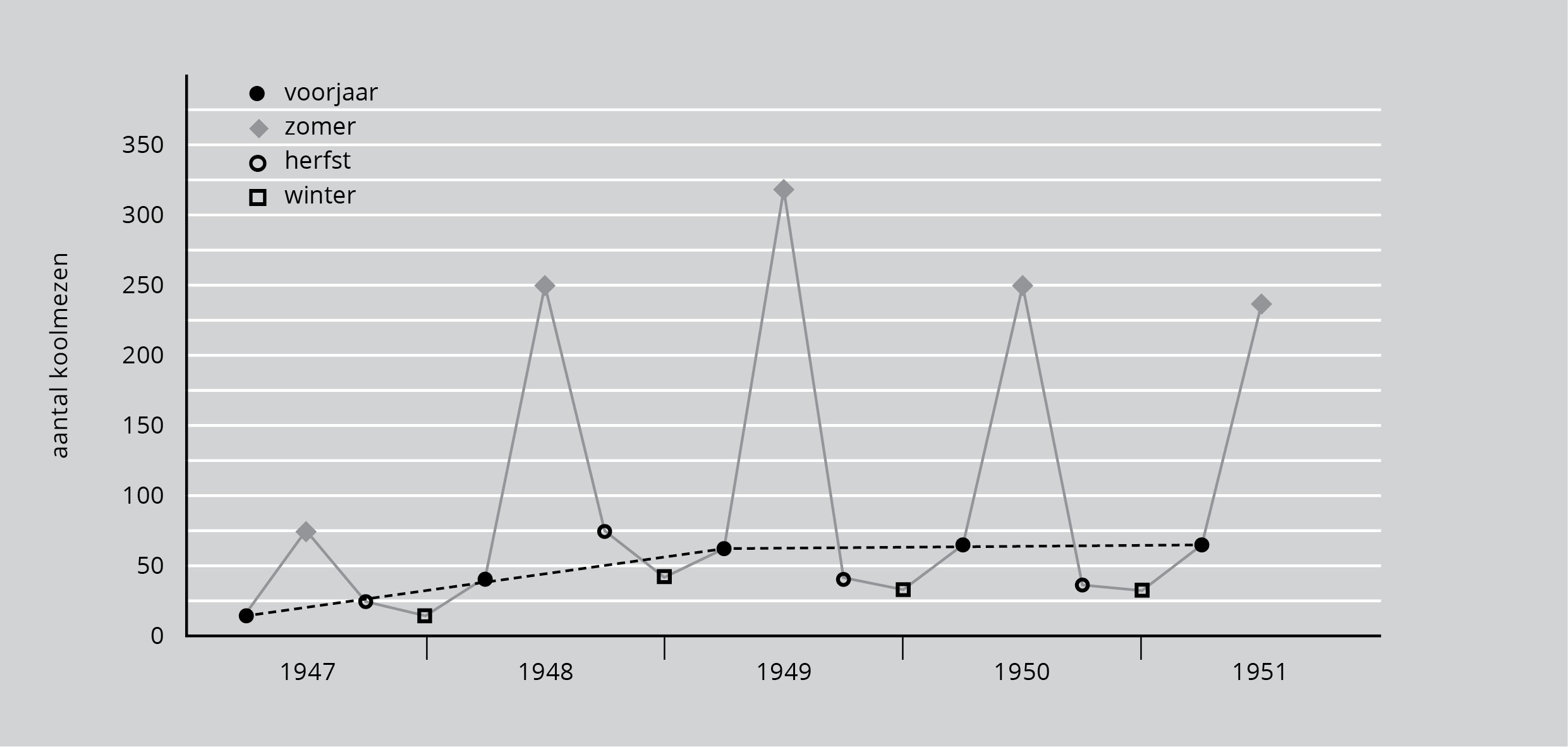
Figuur 6: Veranderingen van een populatie koolmezen per seizoen en per jaar (bron: D. Lack, 1954).
De populatie begon klein. Misschien kwam dit door de voorafgaande strenge winter, of doordat steeds meer vogels de nestkastjes gebruikten. Niet alle jonge koolmezen die in de herfst verdwijnen, hoeven te zijn gestorven, maar het feit dat de meerderheid niet in het bos is gebleven, doet vermoeden dat de draagkracht van het lokale milieu al door de overblijvers wordt bereikt. Al is de groei elke keer weer enorm, door de jaren heen heeft dit geen invloed op de koolmezenpopulatie in het bos. Dit is de normale gang van zaken in de meeste populaties: het groeivermogen is enorm, toch blijft de populatie min of meer stabiel.
Het groeivermogen is enorm, de rem ook.

.
Het valt niet mee om te begrijpen hoe een populatie in de natuur wordt gereguleerd. Dit geldt zowel voor populaties van planten als van dieren. Laten we eens kijken hoe het is bij de reiger. Want er is veel bekend over reigers, al blijven er vragen.
We bekijken de Europese blauwe reiger (onze gewone reiger) en daarnaast ook de Amerikaanse grote blauwe reiger. Deze soorten verschillen in een aantal onbelangrijke details in vorm en pluimage, maar ze komen zó overeen, dat we ze zonder meer als gelijken kunnen behandelen. Blauwe reigers komen hier algemeen voor. Ze voeden zich in hoofdzaak met vis, die ze met hun bek in ondiep water vangen. Daarnaast eten ze ook kleine zoogdieren, watersalamanders, jonge vogeltjes, krabben, garnalen en een allerlei soorten waterinsecten. Bijna alles in hun leven is gericht op het vangen van vis. Omdat reigers niet naar voedsel kunnen duiken en zelden zwemmen, moet het water waarin ze vissen ondiep genoeg zijn om te kunnen waden en helder genoeg om de vis te kunnen zien.
Reigers broeden in kolonies. Die bestaan soms uit honderd of meer grote nesten. Veel kolonies zijn veel kleiner en ook broeden paren wel apart. Ieder jaar worden weer dezelfde nestplaatsen gebruikt. De nesten bevinden zich tot meer dan twintig meter hoog in de bomen, maar in gebieden waar weinig bomen zijn, worden de nesten soms op de grond gemaakt. Reigerkolonies liggen zo gunstig mogelijk ten opzichte van de viswaters, zodat deze voedselgebieden optimaal benut worden.
Zo’n keuze van een centraal gelegen plaats is normaal voor dieren. Het bestaan ervan doet ecologen veronderstellen dat vogelpopulaties in hun omvang vooral beperkt worden door de beschikbaarheid van voedsel.
Ook planten staan meestal zo opgesteld dat ze optimaal profiteren van de omstandigheden.
Bij de reiger vormt de keuze van een centrale plaats voor het broedgebied in elk geval een aanwijzing voor het belang van goede visgronden tijdens het broedseizoen. Goede visgronden worden gekenmerkt door de ruime beschikbaarheid van voedsel. Er moet onderscheid worden gemaakt tussen rijk voorkomend en beschikbaar, bereikbaar voedsel. Vis op grote diepte is onbereikbaar voor de reiger. Hij heeft er niets aan, al is er nog zoveel van. Dit verschil tussen rijk voorkomend en beschikbaar voedsel komen we vaak tegen. Het onderscheid is van belang bij de bestudering van de rol van voedsel bij de regulering van de omvang van populaties.
Het aantal zilverreigers, in 1980 nog een dwaalgast, begint het aantal blauwe reigers te naderen. Maar verdringing is het niet.

.
Op twee plaatsen in Engeland hebben vogelwachters jarenlang tellingen uitgevoerd van zoveel mogelijk nesten. We kunnen er van uitgaan dat wat voor Engeland geldt, ook voor Nederland opgaat. De nesten zijn in het late voorjaar geteld op het moment dat de meeste paren jongen hadden. In die tijd van het jaar is het mogelijk vanaf de grond te beoordelen welke nesten bewoond worden en welke niet. Bewoonde nesten hebben verse groene twijgen langs de rand en er ligt duidelijk zichtbare ‘witkalk’ onder, veroorzaakt door de vloeibare uitwerpselen van de jongen.
Een reigerpaar brengt in de meeste jaren met succes drie jongen voort. Voor een kolonie van 300 paren betekent dit dat er na het broedseizoen 900 reigers bij komen, wat het aantal op 1500 brengt.
Figuur 7 laat zien dat zó’n stijging zich nooit voordoet. Daaruit volgt dat voor het volgende broedseizoen veel reigers gestorven of uit het gebied verdwenen zijn. De twee lijnen tonen wel dat er van jaar tot jaar grote schommelingen in het aantal nesten optreden. Maar deze schommelingen in de populatie zijn dus klein vergeleken bij wat theorétisch mogelijk is.
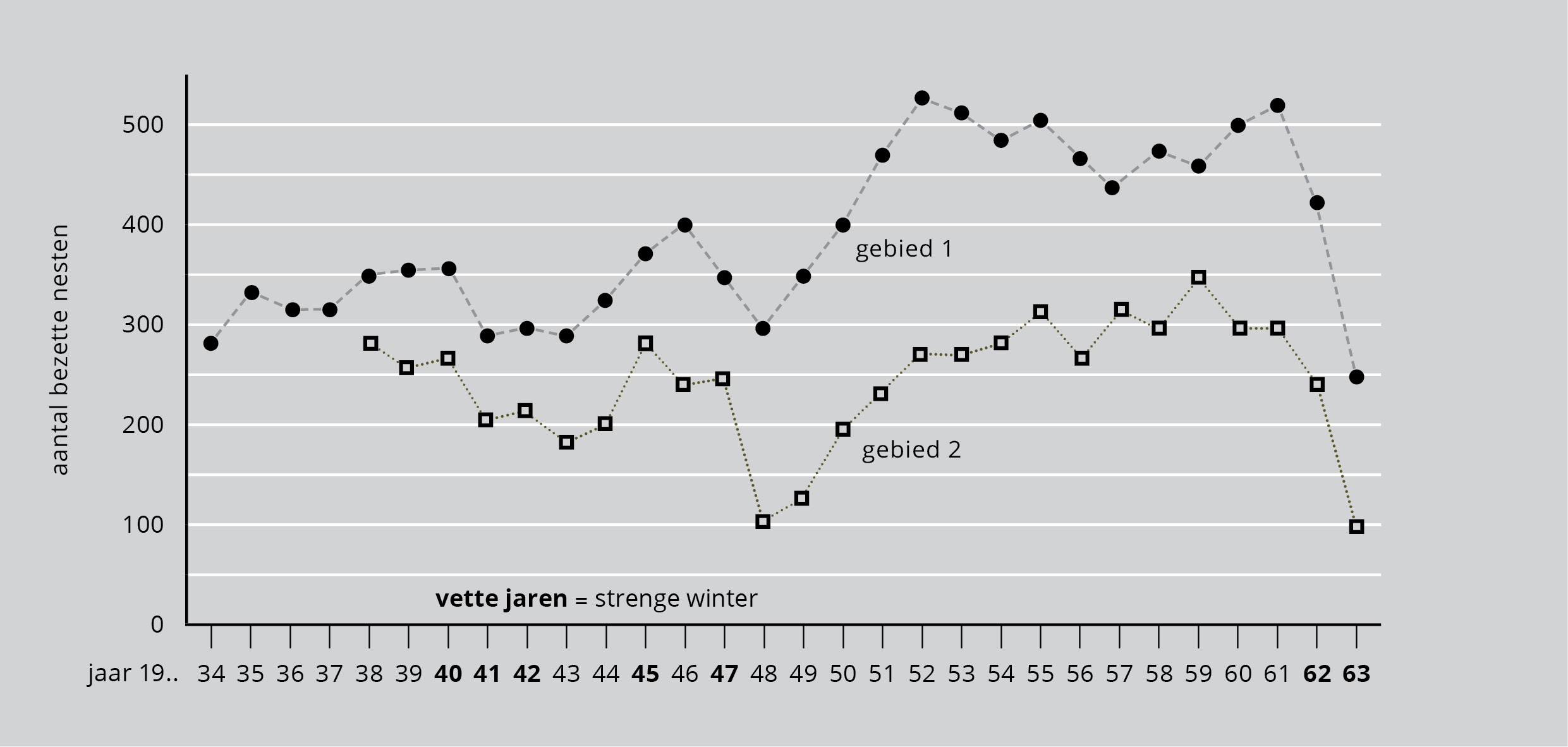
Figuur 7: Aantal broedende reigerparen in twee gebieden in Engeland (Thameswaterschap, resp. Cheshire en South Lancashire) (bron: D. Lack, 1966).
In Engeland blijven veel reigers overwinteren. Een vogel die afhankelijk is van voedsel uit ondiep water, zal gauw gebrek lijden als het water bevriest. De broedende populatie is na een strenge winter dan ook meestal kleiner, maar het kost slechts enkele jaren om weer op het oude peil te komen. De populaties in figuur 7 vertonen beide dezelfde schommelingen: een slecht jaar in het ene gebied is ook slecht in het andere, waardoor de gevolgen van strenge winters nog benadrukt worden.
De grootste kolonie bij Nieuwkoop had op zijn toppunt in 1975 439 nesten. Zie ook dit kaartje.

.
Door reigers jarenlang te ringen was het mogelijk om de trek en de sterfte na te gaan. Op deze genummerde ringen staat het verzoek aan ieder die een vogel vindt, dit te melden aan het vogeltrekstation. Het aantal meldingen van geringde vogels hangt af van de soort. Er komen betrekkelijk weinig meldingen binnen van kleine soorten of soorten die naar veraf gelegen gebieden trekken. Maar van grote vogels zoals reigers is het aantal meldingen hoog. Er zijn veel reigers geringd en het aantal meldingen is voldoende om een schatting te kunnen maken van het sterftecijfer.
Figuur 8 laat de sterfte van jonge Amerikaanse reigers zien vanaf het moment dat ze het nest verlaten hebben. Het toont van elke maand de sterfte (te beginnen in juli) en geeft daarbij de leeftijd in maanden van de jonge reigers aan. We kunnen zien dat veel jonge vogels sterven tijdens de zomer dat ze het nest verlaten. Het sterftecijfer blijft hoog tot de maand februari van het volgende jaar. Daarna neemt het sterftecijfer sterk af.
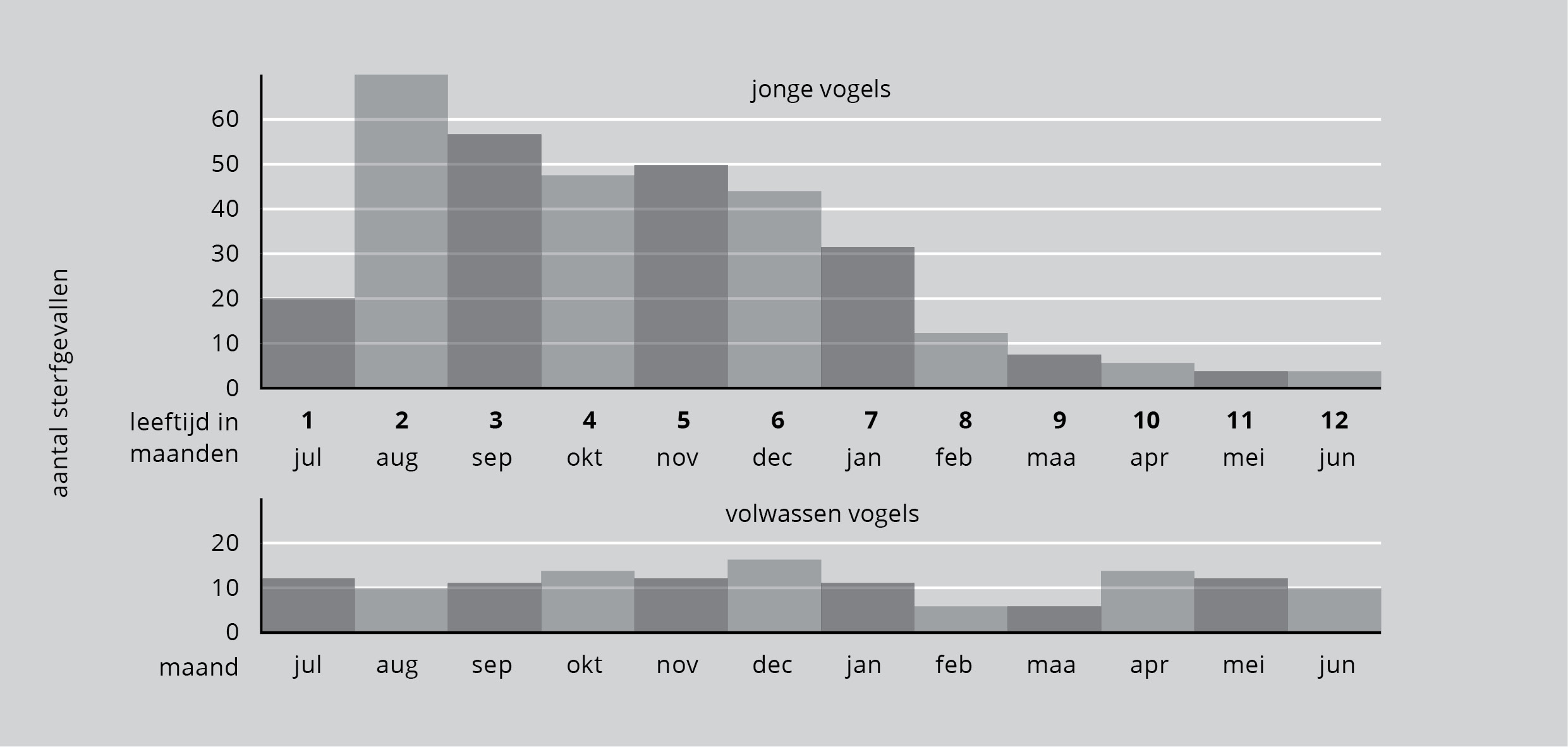
Figuur 8: Boven: het aantal teruggevonden geringde vogels van nog geen jaar oud, per maand.
Veel reigers sterven dus als ze nog maar een paar maanden oud zijn, mogelijk vanwege onervarenheid in het vangen van vis. Voor de volwassen vogels is de kans te sterven alle seizoenen ongeveer gelijk. Klaarblijkelijk vormen de eerste zes maanden de kritieke periode voor de reiger. Daarna verbeteren de kansen op overleving sterk.
De cijfers zijn zelfs nog dramatischer als de sterfte wordt afgezet naar de leeftijd zoals in figuur 9. De overlevingskans neemt na het eerste levensjaar sterk toe en tegen de tijd dat de vogels drie jaar zijn geworden en beginnen met broeden is de kans te sterven klein geworden. Vanaf dat moment is het sterftecijfer laag.
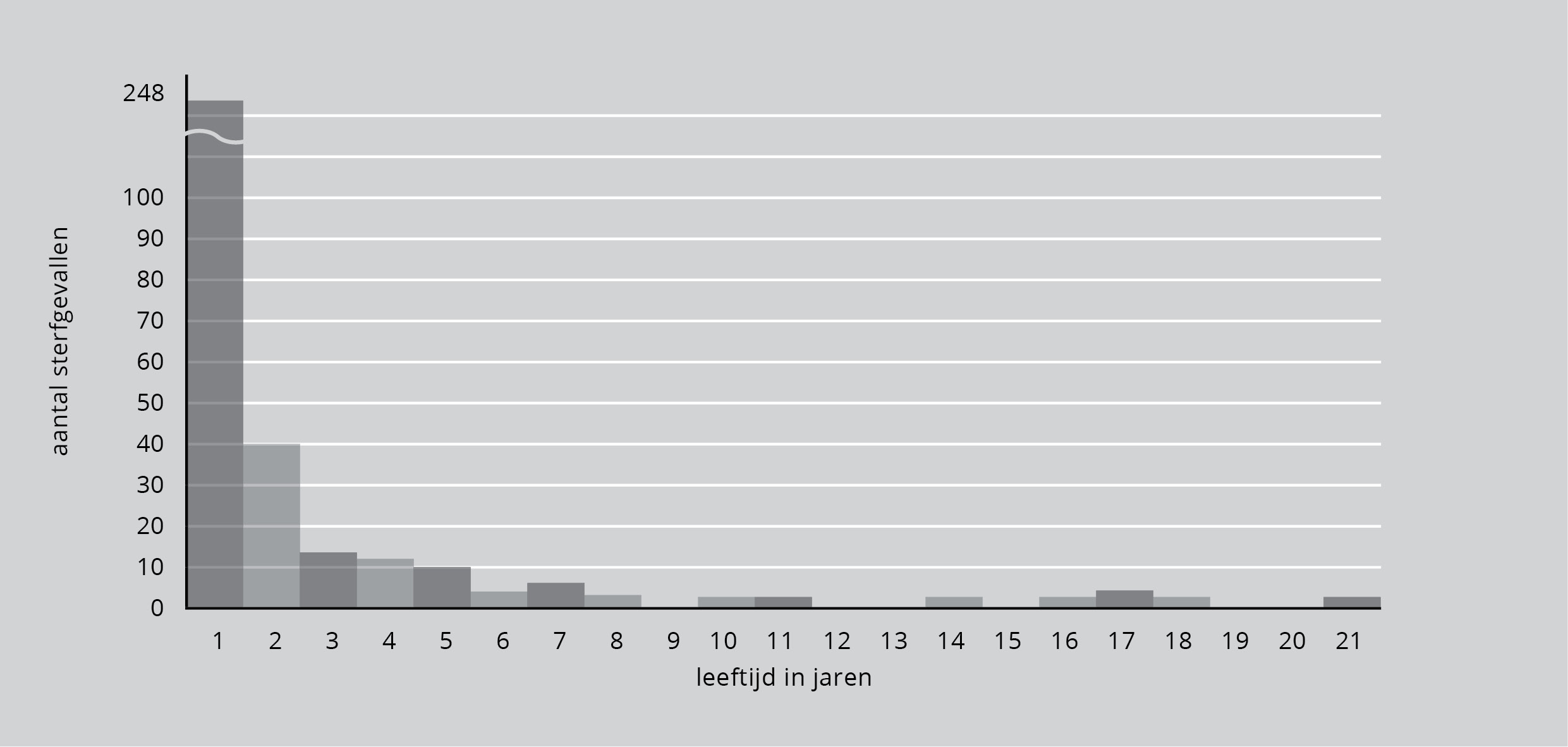
Figuur 9: Het sterftecijfer van de Noord-Amerikaanse grote blauwe reiger afgezet tegen de leeftijd.
Een vergelijking van cijfers voor heel Noord-Amerika en voor het kleinere onderzoeksgebied in Engeland levert bijna identieke uitkomsten op: een hoge sterfte in het eerste jaar van gemiddeld 70 procent.
Het hoge sterftecijfer voor jonge vogels is kenmerkend voor praktisch alle andere vogelsoorten die op deze manier zijn bestudeerd. Wijzelf zijn gewend om sterfte met ouderdom te verbinden en het is misschien een ontnuchterende gedachte dat de meeste vogels die in je tuin geboren worden, sterven voor ze één jaar oud zijn en voor ze de kans krijgen te nestelen.
In een strenge winter staat de dode blauwe reiger soms vastgevroren in zijn eigen sloot.

.
Reigers leggen hun eieren met tussenpozen van twee dagen. De omvang van het nest varieert van twee tot vijf eieren. In zeldzame gevallen zijn het er zes. Het gemiddelde is vier.
Reigers broeden meestal niet voor ze twee jaar oud zijn. Het broeden begint als het tweede of derde ei gelegd is. Zodoende komen de eerste twee of drie eieren op dezelfde dag uit en er zijn gewoonlijk steeds tussenpozen van ongeveer twee dagen voordat de daaropvolgende eieren uitkomen.
Omdat de jongen snel groeien is er dus verschil tussen de jongen die het eerst uitkomen en de latere. Heel vaak is het laatste jong buitengewoon klein vergeleken met zijn broertjes en zusjes.
Dit om de beurt uitkomen komt vooral voor bij de grote roofvogels en is minder gewoon voor kleine zangvogels (zoals de koolmees). De jongen daarvan komen bijna tegelijk uit. We komen op de betekenis hiervan terug. Laten we nu eerst het voedsel van de jonge reigers in het nest bespreken.
De jongen worden door beide ouders gevoed. De ouders vangen daartoe vis in soms afgelegen water zodat ze met tussenpozen naar de kolonie terugkomen met hun maag vol voedsel. Bij het inslikken gaat de kop van de gevangen vis het eerst naar binnen. Op het nest worden ze met de staart naar voren uitgebraakt, omgedraaid en aan de jongen gevoerd met de kop eerst, waardoor de jongen stekelige vis als baars veel gemakkelijker kunnen doorslikken. Als een volwassen vogel langdurig op vangst uit is geweest, is het voedsel dat hij meebrengt al gedeeltelijk verteerd. Als hij snel terugkeert, is het voedsel nog nauwelijks verteerd.
Zoals veel andere visetende vogels spuwen de jongen het voedsel uit als ze door een indringer gestoord worden. Dat doen ze vermoedelijk om de aandacht van potentiële roofdieren af te leiden. Wie naar een nest met jongen klimt, krijgt de volle laag van uitgebraakte vis! Van dit gedrag kan gebruik gemaakt worden om uit te vinden wat ze precies eten. Veel van het voedsel is gemakkelijk thuis te brengen en kan daarom direct in het nest worden teruggelegd, zodat de jongen het opnieuw kunnen eten.
Reigers brengen minder succesvolle jongen voort dan ze eieren leggen.

.
Het voedsel van reigers is zeer gevarieerd. Per woongebied bestaan er opvallende verschillen in de voedselsamenstelling van de jongen. In een bepaald gebied overheerst bijvoorbeeld witvis als voedsel, terwijl in andere gebieden paling of stekelbaarsjes, maar ook kikkers of zelfs garnalen de hoofdmaaltijd kunnen vormen. Deze verschillen hangen samen met de verschillende milieus van de jagende volwassenen. In de Betuwe bijvoorbeeld jagen ze vooral langs de oevers van de (grote) rivieren. Bij Nieuwkoop vinden de vogels hun voedsel in de plassen en in de sloten van de omliggende polders. Langs de kust wordt ook wel in brak water gejaagd, wat de vangst van garnalen verklaart.
Zo nu en dan vangt de reiger een mol of een muis (vooral waterspitsmuizen). Soms wordt er een ander zoogdier gevangen, tot zelfs een volwassen hermelijn die toch zelf een venijnige jager is. Insecten schijnen meer toevallig meegepikt te worden als de reigers op een grotere prooi staan te wachten. De meeste gevangen waterjuffers zijn net uitgekomen exemplaren, die nog te jong zijn om goed te kunnen vliegen en de waterkevers die de reiger vangt zijn de grotere soorten.
Volwassen reigers vangen natuurlijk niet alle vis die ze aantreffen. Vissen van meer dan 25 cm zijn te groot. De meeste gevangen vissen, blankvoorn, baars, snoek en dergelijke soorten, zijn tussen 11 en 20 cm lang. Dat is dus kleiner dan waar de gemiddelde sportvisser belangstelling voor heeft. Prooidieren van minder dan 10 cm lang worden meestal ook met rust gelaten, terwijl ze toch talrijk moeten zijn op de plaatsen waar de reigers jagen. Stekelbaarsjes en garnalen vormen een uitzondering en worden misschien alleen gevangen wanneer grotere dieren niet beschikbaar zijn.
Uit zijn dieet kan men concluderen dat de reiger alles pakt wat hij redelijkerwijs aan kan, maar zich het liefste concentreert op vis van gemiddelde grootte. Men mag aannemen dat de vangst van dag tot dag verschilt, en dat het voedsel ook per seizoen en per jaar wisselt. Deze veranderingen in de beschikbaarheid van voedsel zullen het groeitempo en de overlevingskans beïnvloeden.
In juni en juli beginnen de jongen de kolonie te verlaten. Dan schijnt de beschikbaarheid van het voedsel te verminderen als gevolg van de plantengroei langs de oevers van plassen en meren. De vangst in de sloten is later in het seizoen soms zelfs bijna uitgesloten door de dichte groei van waterplanten aan het oppervlak. De reiger vangt het meest bij droog weer, wellicht omdat bij regen het water modderig wordt. Deze factoren bepalen mede het vermogen van de volwassenen om hun jongen met succes groot te brengen.
De jongen verlaten de kolonie dus na een paar maanden.

.
We krijgen een idee van de veranderingen in de voedingsomstandigheden door de jongen periodiek te wegen. Men gebruikt bij vogels en zoogdieren de lichaamsgewichtstoename vaak als maatstaf voor de hoeveelheid opgenomen voedsel. Figuur 10 laat de veranderingen in het gewicht van de vijf jongen uit één nest zien vanaf het moment dat ze uit het ei komen totdat ze bijna volgroeid zijn. Twee eieren komen uit op 2 mei, een derde op 3 mei, een vierde op 5 mei en het laatste op 8 mei.
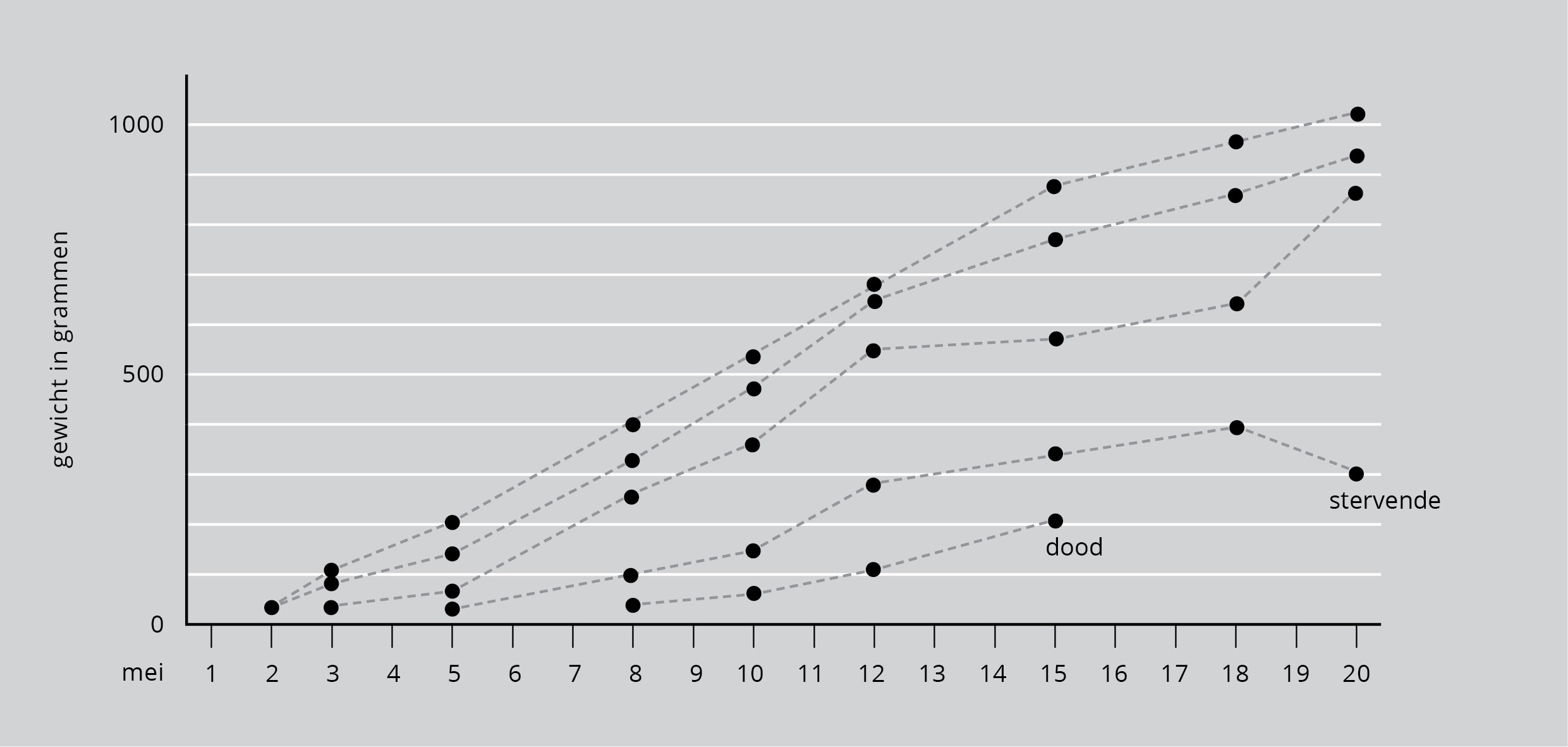
Figuur 10: Veranderingen in het gewicht in een nest van vijf blauwe reigers.
De eerste twee jongen groeiden snel. Het derde ook, tot 12 mei. Toen begon de groei van nummer drie af te nemen. Het vierde jong groeide langzamer en stierf door gebrek aan voldoende voedsel op 20 mei. Bijna direct daarop kwam het derde jong weer aan en werd al spoedig weer bijna even zwaar als de eerste twee. Dat duidt erop dat de aanwezigheid van het vierde jong de groei van het derde remde. De laatst uitgekomen, vijfde vogel leefde maar een week en had vanaf het begin geen kans.
Deze gebeurtenissen tonen wat er in een reigernest gebeurt als er een groot gezin gevoed moet worden in een slecht jaar. In zulke jaren slagen broedsels van twee of drie jongen meestal volledig, maar van broedsels van vier of vijf jongen sterven er één of twee van de honger. Van tijd tot tijd gebeurt het dat grote broedsels feitelijk minder succes hebben dan kleine, want daar moet het voedsel over meer jongen worden verdeeld. Maar in goede jaren, als het weer droog is en de visvangst beter, hebben de grotere broedsels wel succes en veel paren kunnen dan vier of zelfs vijf jongen groot brengen.
We kunnen nu het voordeel zien van het niet tegelijk uitkomen van de eieren. Als alle eieren tegelijk uitkwamen en de jongen tijdens de groei even groot waren, zou de onderlinge concurrentie kunnen leiden tot verlies van het hele nest. Maar als de eieren niet tegelijk uitkomen, zijn er altijd kleine jongen bij. Ze zijn zwakkere concurrenten wanneer hun ouders met voedsel terugkeren en ze overleven alleen als er veel voedsel is. De eerstgeboren jongen overleven bijna altijd en vliegen uit.
Wat vertellen de vondsten van dode, geringde vogels in juli en augustus ons, toen die dus pas een paar maanden oud waren? Ze laten zien dat na het verlaten van het nest, de kans om te overleven voor een deel afhangt van de omvang van het broedsel: vogels uit grote nesten hebben wat minder kans om te overleven dan die uit kleine nesten. Vermoedelijk komt dit, doordat in middelmatige tot goede jaren de jongen van grote nesten allemaal uitvliegen en ze in iets minder goede vorm verkeren en iets minder wegen dan de exemplaren van kleine broedsels. Het verschil is niet groot, maar het is voldoende om te laten zien dat grote broedsels, die in het nest succesvol leken, het later niet altijd even goed doen.
Met meerdere jongen per nest is de populatie weerbaar tegen slechte jaren.

.
Reigers leven min of meer op zichzelf, behalve als ze in het vroege voorjaar samenkomen om op traditionele plaatsen te broeden. De omvang van de kolonie hangt af van de draagkracht van het plaatselijk milieu, dat hoofdzakelijk bepaald wordt door de beschikbaarheid van vis. De beschikbaarheid van voedsel is van meer belang dan de rijkdom eraan, want als het voor de reiger onbereikbaar is, heeft hij er niets aan. Het belang van de beschikbaarheid is treffend en moet gezien worden tegen de achtergrond van de vangstmethode van de reiger. Daarbij spelen behalve de diepte van het water, ook weersomstandigheden en jaargetijde een rol.
Al deze omstandigheden en de bekwaamheden van hun ouders bepalen de overlevingskansen van de jonge reigers. In magere jaren sterven veel jongen, vooral uit de grote broedsels. Als de vogel het eerste jaar heeft overleefd, heeft hij goede vooruitzichten om zelf verschillende keren jongen voort te brengen. Schommelingen in het aantal reigers van een kolonie houden verband met strenge winters e.d. Het lijkt er op dat, voorzover het reigers betreft, de draagkracht van het milieu geheel kan worden begrepen in termen van de beschikbaarheid van voedsel.
Maar waarom zou, als dat zo is, een in wezen op zichzelf levende vogel, zich sociaal gaan gedragen en voor een klein deel van het jaar in kolonies gaan broeden? Zou verspreid nestelen niet gunstiger zijn, omdat de beschikbare voedselbronnen dan efficiënter verdeeld en benut worden? Het broeden in kolonies moet bepaalde voordelen bieden.
Misschien heeft het te maken met gebrek aan veilige nestplaatsen, of misschien is het omdat de vogels een zekere wederzijdse aanmoediging in het vroege broedseizoen nodig hebben. Reigers vertonen dan een uitgesproken baltsgedrag en steeds opnieuw kan men zien dat een baltsend paar andere reigers stimuleert. Op welke manier een reiger, die voor het eerst gaat broeden, zich in een bestaande kolonie een plaats verovert, is een andere vraag. Vogels van een jaar oud bezoeken de kolonies en bouwen soms kleine nesten, maar ze doen slechts in zeldzame gevallen pogingen te gaan broeden; nooit met succes. Ze schijnen wel deel te hebben in de aangelegenheden van oudere vogels. Ze doen mee met een deel van het baltsgedrag en vaak volgen ze volwassenen die op voedsel uitgaan. Het is bijna alsof er een soort leertijd is, waarin de jonge vogels de oudere imiteren en met vallen en opstaan ervaring in het leven in een kolonie opdoen.
Tenslotte is er de vraag waarom na vele generaties een kolonie plotseling verlaten wordt en hoe nieuwe kolonies begonnen worden. Het is duidelijk dat er nog veel valt te leren over de ecologie van de reiger.
Tot zover wat we kunnen leren van één soort.

.
Uit diverse onderzoekingen is gebleken dat populaties in het algemeen stabiel zijn. Veranderingen blijven binnen de perken door dichtheidsafhankelijke factoren. We bespraken daar voorbeelden van bij de koolmees en de reiger. Stabiliteit van de populatie lijkt regel te zijn, maar er komen ook uitzonderingen voor.
Zo kan een planten- of dierpopulatie een tijdelijke exponentiële groei doormaken, nadat ze door de mens per ongeluk of bewust in een nieuw gebied is geïntroduceerd. Er bestaan ook enkele natuurlijke uitzonderingen. Deze komen voornamelijk voor onder dieren die op hoge breedtegraad leven (bij de polen), of in droge gebieden gelegen in of bij de tropen. In deze gebieden kunnen de schommelingen van de aantallen een omvang bereiken die men elders zelden aantreft.
De beide poolgebieden worden gekenmerkt door een klein aantal soorten planten en dieren, die dan wel in gróte aantallen voorkomen. Iets dergelijks vinden we in woestijn- en steppegebieden, hoewel hier ook flink wat exemplaren van weinig voorkomende soorten aanwezig kunnen zijn. Wie ’s zomers naar IJsland gaat, raakt evenzeer onder de indruk van de grote aantallen van sommige zeevogels, nachtvlinders en muggen, als van het ontbreken van dagvlinders en de meeste zangvogelsoorten. Op de droge savanne in Afrika treft men grote aantallen van enkele soorten wevervogels en witte vlinders aan, maar veel dieren van de vochtiger tropen zal men er niét vinden. De populatie van bepaalde soorten wordt in zulke milieus instabiel.
De instabiliteit kan zich op twee manieren manifesteren, namelijk als regelmatige schommelingen in de omvang van een populatie of als incidentele toename.
Bij regelmatige schommelingen ziet men na een periode van geringe aantallen een aanzienlijke toename. Die vindt plaats binnen een regelmatig en voorspelbaar tijdsverloop. Het proces zet zich jaar in, jaar uit voort en de schommelingen zijn zo regelmatig dat ze wel populatiekringlopen genoemd worden. Hoewel ecologen het eens zijn over het bestaan van de populatieschommelingen, vechten sommigen de beweerde regelmaat aan, terwijl anderen deze misschien overdrijven. Voor enkele soorten schijnen regelmatige populatieschommelingen in elk geval een feit. Als dit het geval is bij prooidieren, kan soms ook de populatie van het roofdier schommelen.
Van soorten waarbij een incidentéle toename van de populatie voorkomt, is het tijdstip en de grootte van de toename niet te voorspellen. Als de aantallen toenemen, ontstaat er bij mobiele dieren, zoals vogels en gevleugelde insecten, vaak een omvangrijke trek. Een dergelijke trek wordt uitbarsting genoemd, of ook wel invasie in het gebied waarheen de uitbarsting zich richt. Men noemt zo’n uitbarsting een plaag wanneer de organismen schade toebrengen aan de menselijke gezondheid of aan de oogst.*
Wat voor toename is de groei van het aantal luizen in het voorjaar?

.
De bekendste regelmatige populatiekringloop is die van de lemming. De lemming is een klein knaagdier in de toendragebieden van Eurazië en Noord-Amerika. De populatieschommeling van de lemming heeft elke vier jaar een piek en een dieptepunt. Als de piek optreedt ondernemen de lemmingen soms lange tochten, vermoedelijk omdat het plaatselijk milieu hen niet langer kan onderhouden. Verhalen over lemmingen die zich van de steile rotswand in zee storten, zijn overigens waarschijnlijk overdreven. De piek in het aantal lemmingen wordt gevolgd door een ineenstorting van de populatie. Daarna kunnen ze zo schaars zijn, dat je ze amper kunt vinden.
De populatiekringloop van de lemming wordt op de voet gevolgd door een kringloop in de populaties van zijn jagers, zoals skua’s (een valkachtige zeevogel) en uilen. Het is niet zozeer dat het aantal jagers schommelt. Deze trekken veeleer de gebieden in waar veel lemmingen zijn. Ze scholen daar samen zolang er veel lemmingen zijn en verspreiden zich als ze weer schaars worden.
Er bestaat geen bewijs dat er zoveel jagers van de lemming leven dat die verantwoordelijk kunnen zijn voor de ineenstorting van hun populatie. Lemmingen zijn planteneters en het is heel wat waarschijnlijker dat de ineenstorting wordt veroorzaakt door voedselgebrek. Die treedt op wanneer het milieu zodanig is vernield, dat verdere toename van de aantallen onmogelijk wordt.
We moeten echter toegeven dat het helemaal niet zeker is dat grote aantallen lemmingen van gebrek omkomen en dat daardoor de populatie afneemt. Maar het schijnt dat het plantenleven rijker wordt, wanneer de toendra tijdelijk bevrijd is van de druk van de lemmingen. Dit geeft de lemmingen dan weer de gelegenheid om in aantal toe te nemen en zo gaat het proces door.
Er is ook wel gesuggereerd dat de populatieschommeling van de lemming wordt veroorzaakt door een inwendige fysiologische kringloop onder de individuen die de populatie vormen. Verschillende soorten knaagdieren in de poolstreken kennen zonder twijfel veranderingen in hun fysiologie die samenhangen met hun aantallen. Het is echter niet duidelijk of deze veranderingen de kringlopen veroorzaken of dat de kringlopen de fysiologie beïnvloeden.
Lemmingen zijn dieren uit de poolstreken. Men vraagt zich af of ze ook populatiekringlopen zouden kennen als ze dichterbij de evenaar leefden. Met andere woorden, maakt het relatief ongecompliceerde milieu van de Noordpool hun kringlopen mogelijk, of heeft het iets met de lemmingen zelf te maken? Zo blijven er altijd vragen.
De verhouding tussen de aantallen jagers en prooien is in de gematigde streken meestal wel ondubbelzinnig.

.
Spectaculair zijn de sprinkhanenplagen. Deze horen bij de droge gebieden in de tropen en de subtropen. De meeste sprinkhanen* eten levende planten, met name grassen, en ze komen dan ook vooral voor in de grasgebieden van de savannes. Sprinkhanen kunnen enorm veel eten. Ze eten per dag hun eigen gewicht aan voedsel op.
Bijna alle sprinkhanenpopulaties blijven normaal tamelijk stabiel, net als die van andere organismen. Maar onder bepaalde omstandigheden kunnen de aantallen van sommige soorten toenemen tot de omvang van een plaag. Ze kunnen dan over grote afstanden trekken, waarbij ze enorme schade toebrengen aan oogsten en andere plantengroei.
In Afrika bestaan drie soorten sprinkhanen die ernstige plagen kunnen vormen en we bekijken hier één daarvan: de woestijnsprinkhaan.
De woestijnsprinkhaan komt voor in het hele gebied van de drogere savannes en woestijnen van Afrika ten zuiden van de Sahara, en verder in Arabië en in Pakistan. Bij lage dichtheid gedraagt de populatie zich als die van de talloze andere soorten sprinkhanen, maar periodiek stijgt hun aantal snel. Dit valt vaak samen met regenval buiten het seizoen en de daarop volgende plantengroei op droge plaatsen. De Rode Zeekust staat reeds lang bekend als bakermat van massale uitbarstingen van de woestijnsprinkhaan. Al duizenden jaren vrezen de boeren in Egypte hun komst.
Veranderingen in het aantal woestijnsprinkhanen hebben te maken met veranderingen in de hoedanigheid van de populatie. Als de populatiedichtheid laag is, verkeren de sprinkhanen in een toestand die bekend staat als de individuele fase. Maar als hun aantallen toenemen, gaan ze over naar wat men de groepsfase noemt. De sleutel tot het begrijpen van het ontstaan van sprinkhanenplagen is het antwoord op de vraag: waarom gaan de sprinkhanen ineens over van de ene fase in de andere?
Nu het klimaat in de war is, zal uitzonderlijke regenval vaker voorkomen.

.
Exemplaren van sprinkhanen in de individuele en in de groepsfase verschillen in hun gedrag èn in uiterlijke kenmerken, zoals hun kleuring. De individuen in de groepsfase hebben de neiging zich samen te voegen: hun gedrag is sterk op elkaar af gestemd. Bovendien zijn ze mobieler en de ontwikkeling van het ei tot volwassen exemplaar verloopt sneller. De onvolwassen sprinkhanen uit deze fase zijn zwart en geel, terwijl die in de individuele fase groen zijn. Sprinkhanen in de groepsfase zijn actiever en socialer dan die in de individuele fase. Ze buiten de omstandigheden uit en hun vermogen tot zwermen is groter dan dat van sprinkhanen in de individuele fase.
De sprinkhanenpopulatie gaat over van individuele naar groepssamenstelling onder invloed van een stijgende populatiedichtheid. Deze dichtheid is sterk afhankelijk van klimatologische omstandigheden. Plotselinge regenval of een overstroming doet de planten snel opschieten en dit veroorzaakt een plaatselijk sterk toenemen van het aantal sprinkhanen. De daarop volgende droogte doet de lokale vegetatie weer sterk inkrimpen. De sprinkhanenpopulatie wordt dan gedwongen nieuwe voedselgebieden te gaan opzoeken. Als de sprinkhanen nog in het onvolwassen stadium verkeren, waarin ze nog niet kunnen vliegen, marcheren ze in lange rijen in één bepaalde richting tot ze meer voedsel tegenkomen.
Daar hopen ze opeen. Hun aantal neemt snel verder toe en het proces herhaalt zich. Eenmaal volwassen kunnen ze vliegen en als het voedsel op is ondernemen ze lange vluchten, waarbij ze zich laten meevoeren met de wind. Er komen nieuwe generaties en het proces herhaalt zich verschillende keren op steeds grotere schaal. Binnen een paar jaar kunnen er honderden miljoenen exemplaren zijn. Dan beginnen zwermen sprinkhanen gebieden binnen te trekken, waar ze jarenlang afwezig zijn geweest. Hun aantallen zijn zo toegenomen dat de plantengroei samen met de oogst praktisch weggevaagd kan worden.
Zulke plagen waren in de oudheid reeds bekend: “Zij bedekten de gehele oppervlakte van het land, zodat het land er door verdonkerd werd en zij vraten al het veldgewas af en alle vruchten van de bomen,” staat in de Bijbel.
Een sprinkhanenplaag kan jaren voortduren. Heel plotseling kan de plaag ook weer ophouden. De insecten keren terug naar de individuele fase en nemen hun rol als gewone sprinkhanen weer op zich.
Sprinkhanen van een plaag richten zich met succes naar de omstandigheden en een plaag kan zozeer aan kracht winnen, dat het alles wat groeit over uitgestrekte gebieden vernietigt. Toch is het einde ervan even opmerkelijk en onverwacht als de oorspronkelijke toename. Daar begrijpen we nog lang niet alles van.
Het is opmerkelijk dat zo’n plaag zoveel generaties kan aanhouden.

.
Ook in Nederland kennen we insectensoorten waarvan, net als bij de sprinkhaan, twee typen bestaan en waarbij ook de aantallen opvallend veranderen. De oorzaken zijn echter volkomen verschillend. Iedereen kent het kleine tienstippige lieveheersbeestje (Coccinella undecempunctata) met zijn typische stippeling op de dekschildjes.
Dit lieveheersbeest kent in Nederland twee verschillende vormen: de ene heeft oranje dekschilden met donkere vlekken en de ander bezit donkere dekschilden met een oranje stippeling. Oranje dieren met donkere stippeling worden het meest geboren. De oranje vorm is dominant. De donkere soort is echter veel beter bestand tegen de winterkoude dan de oranje soort. (We noemen deze laatste minder winterhard.) Het aantal exemplaren van de oranje vorm is in het zomerseizoen dus groter. ’s Winters sterven grote aantallen van de oranje vorm, terwijl er veel minder exemplaren van de donkere vorm doodgaan. Dan heeft de donkere vorm een veel belangrijker aandeel in de populatie. In de lente neemt het aantal van de oranje dieren weer toe.
De instabiliteit heeft hier dus de regelmaat van het seizoen. De oorzaak ervan ligt niet in het verband tussen de populatiedichtheid en de voedselsituatie, maar in het klimaat. Van één soort bestaan twee typen, waarvan er één meer weerstand heeft tegen de dichtheidsonafhankelijk werkende factor van het klimaat. Dit veroorzaakt dat elk type zijn eigen schommeling doormaakt.
De groene planten vormen een aparte categorie. Planten hebben ruimte nodig om te groeien en om de stralingsenergie van de zon te vangen. Ze hebben ook water en anorganische stoffen nodig. Ze kunnen zich niet verplaatsen om water- en voedselbronnen te gaan opzoeken. De concurrentie tussen planten richt zich op een geschikte plaats om te groeien. Succes in déze concurrentiestrijd bepaalt de overlevingskans. Er is dan ook alle reden om aan te nemen dat de dichtheidsafhankelijkheid, evenals bij dieren optreedt bij planten. Mensen die een tuin hebben, zijn ermee vertrouwd dat de onkruiden om de ruimte concurreren met hun planten; het wieden van onkruid of het uitdunnen van zaaiplanten is voor tuinier en boer zo gewoon, dat men niet bij de betekenis ervan stil staat.
Er zijn ook onafhankelijk van de dichtheid optredende factoren die de omvang van de plantenpopulatie beïnvloeden. Planten kunnen niet als dieren beschutting zoeken als het weer slecht is. Dit leidt soms tot het sterven van veel planten, zoals in het geval van ernstige, onverwachte droogte of vorst. Deze aantasting treedt onafhankelijk van de dichtheid op en is bij planten belangrijker dan bij dieren.
Er is het individuele belang van één dier, of één plant en er is het collectieve belang om te overleven als soort.

.
We zagen dat bij reigers de draagkracht van het milieu, gedefinieerd als de beschikbaarheid van voedsel, de belangrijkste factor bij de stabiliteit van de populatie lijkt te zijn.
Bij een sprinkhanenplaag blijft het moeilijk te begrijpen hoe normale sprinkhanen kunnen veranderen in doelgerichte dieren die alles kaalvreten en hoe het komt dat een plaag plotseling weer afneemt. Het kan beginnen met een arm milieu dat plotseling meer voedsel gaat bevatten, toch heeft het daarna waarschijnlijk niet te maken met een tekort aan voedsel, want de sprinkhanen kunnen altijd naar nieuwe voedselgebieden trekken.
Bij de verschillende lieveheersbeestjes is het klimaat een bepalende factor. Bij groene planten gaat het er om wie een geschikte plaats om te groeien weet te bemachtigen. Het gaat om ruimte.
Verliezen door roofdieren doen zich dichtheidsafhankelijk voor. Het hoeft niet de hoeveelheid beschikbaar voedsel te zijn die de omvang van een populatie van planteneters beperkt, want het kan ook zijn dat roofdieren de populatie op een lager niveau houden. Maar het zijn waarschijnlijk niet de reigers die de visstand bepalen.
Hoe zit het met het evenwicht tussen planten en planteneters?
In vochtige tropische wouden is er het hele jaar een overvloed aan plantengroei en kaalgevreten plekken zijn ongebruikelijk. Het is mogelijk dat voedsel daar voor plantenetende dieren geen beperking vormt. Toch zijn planteneters er zelden algemeen. Ze worden belaagd door allerlei roofdieren. Al met al doet dit sommige ecologen geloven, dat in het algemeen dieren die van planten leven, niet door de beschikbaarheid van vóedsel beperkt worden. We zullen op dit vraagstuk later terugkomen.
Er is een andere theorie die stelt dat sommige dieren hun eigen dichtheid regelen beneden het niveau dat de draagkracht van het milieu stelt. De theorie gaat uit van de betekenis van het territorium voor het sociale gedrag. Sociaal gedrag beïnvloedt dan de voortplanting. Dit is vooral onderzocht bij allerlei soorten vogels. Zo zouden reigers door de sociale wisselwerking in de kolonie bij hoge dichtheid minder jongen voortbrengen en bij lage dichtheid meer. De theorie is nooit behoorlijk getoetst. De bewijzen ervoor zijn niet erg solide en kunnen even goed verklaard worden als de directe werking van dichtheidsafhankelijke factoren.
Er zijn óók ecologen die dichtheidsafhankelijke factoren afwijzen en daarentegen stellen dat sterfte meestal onafhankelijk van de dichtheid optreedt. Zij beroepen zich op het klimaat als de voornaamste oorzaak voor de schommelingen in de aantallen. Het klimaat houdt de omvang van de populaties zo beperkt dat voedselgebrek voor dit beperkte aantal individuen niet voorkomt. Sterfte als gevolg van voedseltekort en de activiteiten van roofdieren wordt natuurlijk niet ontkend, maar deze ecologen vinden dat dit geen belangrijke rol speelt bij het reguleren van de totále aantallen.
De meeste bewijzen voor deze theorie komen uit het insectenonderzoek. Het lijdt geen twijfel dat insecten vaak zware verliezen lijden als gevolg van het klimaat. Maar dat het klimaat op de lange duur belangrijker is dan de strikt biologische factoren, die dichtheidsafhankelijke sterfte veroorzaken, is niet meer dan een vermoeden. Het laatste woord is er zeker nog niet over gezegd.
De jagers-verzamelaars in de oudheid waren soms zo ‘wijs’ om hun milieu intact te houden, soms zo ‘dom’ dat zij hun prooidieren finaal uitroeiden.
Hier eindigt de eerste serie van Wat is ecologie? In de volgende serie beginnen we met aflevering nummer 31.
